Опубликовано в журнале:
»» 6 / 2001 ПОРАЖЕНИЕ НЕРВНОЙ СИСТЕМЫ РЕТРОВИРУСАМИ: ПАТОЛОГИЧЕСКАЯ РЕАКЦИЯ ХОЗЯИНА ИЛИ НЕЙРОВИРУЛЕНТНОСТЬ КАК РЕЗУЛЬТАТ ДЕЙСТВИЯ ВИРУСНЫХ ГЕНОВ?
Кристофер Пауэр
Лаборатория нейровирологии, Неврологическая исследовательская группа, г. Калгари, провинция Альберта, КанадаTRENDS in Neurosciences, Vol. 24, No. 3, March 2001.
Ретровирусы представляют важную группу РНК-вирусов, которые вызывают ряд болезней нервной системы. Более того, потенциальную угрозу представляют вновь выявленные ретровирусные инфекции нервной системы и некоторые ритровирусные векторв или белки, используемые как переносчики генов. Данная статья описывает различные ретровирусы и их этиологическую роль в развитии болезней нервной системы, или, по другому, нейровирулентностъ. Специфические последовательности в пределах ретровирусных генов могут быть ответственны за нейровирулентностъ. Другой механизм заключается в активации иммунных реакций хозяина нейровирулентны-миретровирусами, что запускает нейропатогенетический каскад, медиаторами которого служат провоспалительные и нейротоксичные молекулы, а заканчивается все смертью нейрона. Таким образом, ретровирусные инфекции нервной системы иллюстрируют молекулярное взаимодействие между определенными вирусными возбудителями инфекций и патологическими реакциями хозяина; результатом такого взаимодействия является нейровирулентностъ.
Значение вирусных инфекций нервной системы все время возрастает, что видно по появлению за последние 5 лет нескольких глобальных нейрови-русных эпидемий [1]. Кроме того, повышается интерес в использовании вирусных векторов для генной экспрессии и для лечения заболеваний головного мозга [2]. Инфекцию считают нейровирусной, если она соответствует следующим критериям: нейроинвазивность, или проникновение вируса в нервную систему; нейротропизм (см. список терминов) или инфицирование вирусами клеток головного мозга, причем селективно инфицируются нейроны (это и называется нейротропизмом); и нейровирулентность или заболевание нервной системы, индуцированное вирусами. Хотя многие ретровирусы удовлетворяют этим критериям, но некоторые ретровирусные свойства усложняют их ней-робиологию, среди этих свойств: 1) предрасположенность к геномным мутациям; 2) способность вызывать как врожденный, так и адаптационный иммунный ответ в нервной системе; 3) способность вызывать патологические изменения за пределами нервной системы.
Структура и цикл жизни ретровирусов
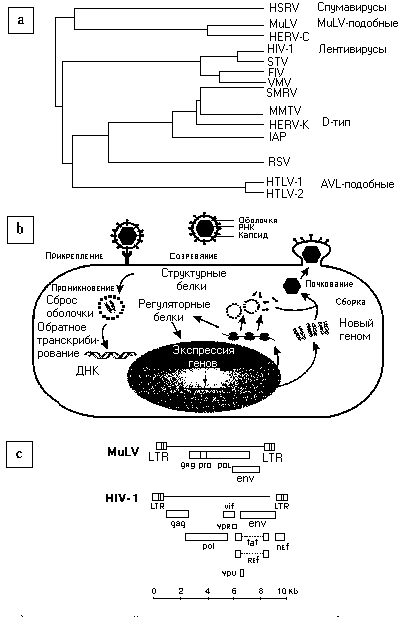
Ретровирусы образуют большую семью оболочечных РНК-вирусов, которые паразитируют на многих видах (рис. 1а) [3]. Эти РНК-вирусы называются ретровирусами, потому что, для них характерна уникальная обратная транскрипция с РНК на вспомогательную ДНК провируса, которая затем интегрируется в хромосомную ДНК хозяина (рис. 1b). После этой интеграции провирус, путем транскрипции, синтезирует вирусные вещества, потенциально способные продлить жизнь клетке, что зависит от факторов транскрипции, продуцируемых клеткой хозяина или вирусом. Все ретровирусы, способные к репликации, имеют три главных открытых фрагментов считывания (в пределах генов gag, pol и env, - см. глоссарий) (рис. 1с). Однако, некоторые группы ретровирусов, такие как лентивирусы, содержат некоторые вспомогательные гены, которые регулируют вирусную репликацию, способность к инфицированию и транскрипцию. Непрочитанный длинный концевой повтор (long terminal repeat - LTR), который располагается на фланге открытых фрагментов считывания и действует как единица промоции, необходим для транскрипции всего вирусного генома и начала транскрипции отдельных вирусных генов. Ретровирус-ный геном представляет не что иное, как линейную молекулу РНК, размером примерно 10 kb [3], которая существует в виде дупликата в интактной вирусной частице или вирионе. Специфические ретровирусные белки, такие, как обратная транскриптаза, интеграза и протеаза кодируются в pol, и они необходимы, соответственно, для вирусной репликации, интеграции и сборки. Ген gag кодирует белки, необходимые для внутриклеточной сборки вируса и его высвобождения из клетки. Ген env кодирует структурные белки (структурная единица поверхности и трансмембранная структурная единица), которые необходимы для связывания с поверхностными молекулами клеток, служащих как рецепторы и медиирующих вход в клетку. У некоторых ретровирусов именно трансмембранные белки ответственны за нейровирулентность, убивая нейроны посредством прямых или косвенных механизмов [4], в последнем случае вначале инфицируются глиальные клетки, из которых затем высвобождаются нейротоксичные молекулы. В качестве непрямого механизма иногда действует активация (и инфицирование) клеток, ответственных за врожденный иммунитет (макрофаги, микроглия, астроциты) или адаптационный иммунитет (Т или В лимфоциты) в нервной системе.
Рис. 1. Ретровирусное филогенетическое дерево, жизненный цикл и репрезентативные структуры генома.
a) На филогенетическом дереве показано несколько нейротропных ретровирусов, в том числе - ретровирусы типа С [вирусы лейкозов грызунов (MuLV) и птиц(АLV), а также вирус, тропный к Т-лимфоцитам человека(НТLV)], лентивирусы [вирусы иммунодефицита человека (ВИЧ-1), обезьян (SIV), кошек (FIV) и вирус visna-maedi (VMV)], спумавирусы и эндогенные ретровирусы человека (HERV-K и HERV-C).
b) Жизненный цикл ретровирусов включает связывание вириона с рецептором клетки хозяина, вход в клетку, затем следует обратное транскрибирование с последующей интеграцией в геном хозяина. Вирусные транскрипты полной длины и отдельные вирусные мРНК синтезируются в ядре клетки, они служат информационным материалом для кодирования как структурных, так и регуляторных белков, которые создают предпосылки для самосборки вирусов и почкования новых вирионов.
c) В качестве наглядного примера представлены геномы ретровируса типа С (MuLV) и лентивируса (ВИЧ-1), показаны важнейшие структурные гены ретровирусов (gag, pol, env), отображена высокая степень сложности генома лентивирусов, обусловленная присутствием дополнительных генов, в том числе, -rev, tat, vif, vpu, nef, vpr.
Иммунологические нарушения, вызываемые некоторыми ретровирусами, приводят ко вторичным инфекциям за пределами нервной системы и к злокачественным процессам в нервной системе. Однако, настоящая статья рассматривает только прямую нейровирулентность ретровирусов, с особым вниманием к механизмам нейровирулентности и ней-ротропизма, кроме того, всесторонне описываются ретровирусы типа С (также именуемые онкогенными), в том числе, вирус лейкемии грызунов (MuLV) и вирусы, тропные к человеческим Т-лимфоцитам (HTLV), а также лентивирусы, такие, как вирусы иммунодефицита человека. Нейротропные
штаммы этих вирусов изучались широко. Они способны вызвать серьезные нарушения здоровья у человека.Ретровирусы типа С (другое название - онкогенные ретровирусы)
Ретровирусы типа С (онкогенные) проявляют нейровирулентность в целом ряде биологических видов, таких, как человек, кошки, грызуны и птицы [4] (таблица 1), что иногда сопровождается параллельным поражением иммунитета. Структура генома этих вирусов проще, чем у лентивирусов, а именно, их геномы включают три структурных гена (два LTR и, в некоторых случаях, имеется добавочный ген). К нейротропным ретровирусам типа С относятся вирусы лейкоза грызунов (MuLV), кошек (FeLV) и птиц (ALV), а также вирусы, тропные к лимфоцитам Т у человека (HTVL-1 и HTVL-2).Таблица 1. Типичные нейротропные ретровирусы: штаммы, нейропатология и клеточный тропизм.
Группа
ретровирусовВирус Патоструктурные измененияв нервной системе а) Клетки нервной системы инфицированные in vivo Тип С CasBrE губчатость, глиоз (СМ) эндотелий, нейроны, глия NE-8 губчатость, глиоз (СМ, СтМ, Мзж) эндотелий, астроциты, нейроны FrCas е) губчатость, глиоз (СМ, СтМ, Мзж) эндотелий, микроглия, нейроны, олигодендроциты Fr98D губчатость, глиоз (Мзж, БВ, СВ) микроглия, нейроны (?) MoMuLV-tsl губчатость, глиоз (СМ, СтМ, Тал, СВ,БВ) эндотелий, астроциты, макрофаги PCV-211 губчатость (Пол, СМ, СтМ, Мзж, БВ) эндотелий TR1.3 кровоизлияние (Мзж) эндотелий LP-BM5 глиоз (Пол, Мзж, БВ) астроциты, микроглия, сосудистое сплетение WB91-GV глиоз олигодендроциты, астроциты HTLV-I/II демилиенизация, глиоз, лимфоцитарнаяинфильтрация (СМ) лимфоциты, астроциты Лентивирусы вирус иммуно-
дефицита человекаглиоз, микроглиальные узелки, многоядерные гигантские клетки, диффузноепобледнение миелина(СМ, Пол, Мзж, БВ, СВ, ПН) макрофаги, микроглия, астроциты вирус visna-maedi глиоз, периваскулярное воспаление, демиелинизация (БВ) макрофаги, микроглия, астроциты вирус иммуно-
дефицита кошекпериваскулярное воспаление, глиоз, микроглиальные узелки (БВ, СВ, Мзж, СМ, ПН) макрофаги, микроглия вирус карпального артрита/энцефалита глиоз, демиелинизация (БВ) макрофаги и микроглия вирус инфекционной
анемии лошадейэпендимит, периваскулрное воспаление (Мзж) клетки периваскулярных зон и мозговых оболочек вирус иммуноде-
фицита коровпериваскулярное воспаление (БВ) глиальные клетки вирус иммуноде-
фицита обезьянпериваскулярное воспаление, глиоз, многоядерные гигантские клетки (БВ, Мзж, СВ, СМ) макрофаги, микроглия, астроциты, эндотелий а) Нейроструктурные изменения даны на основании гистологических заключений. Губчатые изменения и глиоз могут сопровождаться повреждением нервной системы и потерей ее структур, что может быть не заметным при гистопатологиче-ском исследовании.
Сокращения:
СтМ - ствол головного мозга; Мзж - мозжечок; Пол. - большие полушария; БВ - белое вещество; СВ - серое вещество; ПН - периферическая нейропатия; СМ - спинной мозг; Тал - таламус; БВ - белое вещество.Вирусы MuLV
Вирусы MuLV образуют самую большую группу нейротропных ретровирусов. У пораженных грызунов развиваются разнообразные нарушения движений, - чаще всего отмечаются: тремор, паралич задних лап и отсутствие координации [5]. Некоторые штаммы MuLV, попадая в головной мозг, первично инфицируют глиальные клетки, такие, как микроглия, астроциты и олигодендроциты, хотя несколько MuLV инфицируют нейроны. Было выделено несколько рецепторов к MuLV на поверхности клеток [3], в том числе - переносчик катионных аминокислот, тСАТ-1 [6], но до сих пор не известие функционирует ли эта молекула как рецептор цен тральной нервной системы. Считается, что в естест венных условиях MuLV проникают в головно] мозг либо с помощью ими инфицированных лейко цитов, которые перемещаются по нервной системе либо путем инфицирования эндотелиальных кле ток головного мозга. Патологические изменения в нервной системе, вызываемые MuLV, также зависят от конкретного штамма, хотя чаще всего ветречаются "губчатые" изменения или потеря паренхимы ЦНС во всех секторах спинного мозга и ствола.
Следует отметить, что один вид MuLV вызывает геморрагические изменения, подобные инсульту, что отражает его селективный тропизм к эндотелиаль-ным клеткам (таблица 1).Нейропатологические механизмы развития MuLV-инфекции являются примером взаимодействия разных последовательностей в геноме ретровируса, обычно расположенных в генах env или gag, и параллельно действует патологическая реакция организма-хозяина, определяемая генотипом мыши. Например, у штамма Fr-98 MuLV два отдельных участка гена env регулируют реализацию нейровирулентности в особых мышиных линиях [7], при этом нейровирулентность проявляется в разных нейроповеденческих нарушениях, приводящих к смерти. В отличие от этого штамма, его штамм-предшественник, хотя и имеет похожий клеточный тропизм (микроглия), но авирулентен в отношении нервной системы. У некоторых разновидностей MuLV нейровирулентности способствуют изменения во внутриклеточном цикле и в экспрессии белков MuLV, кодируемых геном env, а также изменения уровня вирусной репликации в головном мозге [7]. Патологические реакции хозяина, инициируемые инфекцией MuLV, остаются плохо изученными, но обнаруживалась корреляция между нейро-вирулентностью и повышенным про-воспалитель-ным образованием цитокинов в гиальных клетках [9]. К другим патологическим реакциям хозяина, индуцируемым инфекцией MuLV, относятся изменение проницаемости гематоэнцефалического барьера и повышение внеклеточной концентрации глутамата, что ведет к смерти нейрона посредством механизма внешней цитотоксичности [10].
Проведено картирование некоторых участков генов хозяина, ответственных за резистентность к неврологическому заболеванию, индуцируемому MuLV. К этим участкам относятся FV-1, FV-4 и Akvr-1, кодирующие эндогенные ретровирусные гены env и gag, которые мешают проявлениям нейровирулентности, так как связываются с предполагаемым рецептором к вирусу, или, может быть, блокируют этап после входа [5]. Кроме того, нейровирулентность, вызванная несколькими разновидностями MuLV, зависит от возраста хозяина, причем новорожденные имеют повышенную подверженность, ввиду слабости иммунной защиты.
Вирусы HTVL
HTVL-1 и HTVL-2 - это ретровирусы, которые вызывают неврологические нарушения или неопла-зию у человека, впрочем, одновременно эти два вида нарушений встречаются редко [11]. У небольшой (<1%} доли взрослых больных, инфицированных HTVL-1 или HTVL-2 развивается прогрессирующая миелопатия, характеризующаяся воспалением, особенно выраженным в грудном отделе спинного мозга. Белки и геном вирусов [12] выявлялись в астроцитах и лимфоцитах в зонах поражения спинного мозга, но вот рецептор к вирусу, требующийся для вхождения вирусов в клетку, остается неизвестным. Считается, что подобно MuLV, HTVL-1 или HTVL-2 проникают в нервную систему, путешествуя в лимфоцитах. Определенные гаплотипы человеческого лейкоцитарного антигена коррелировали с повышенной подверженностью к HTVL-индуцирумой миелопатии, которая характеризуется усиленной интратекальной иммунной реакцией к вирусу [13]. Патологические изменения в нервной системе характеризуются лимфоцитарной инфильтрацией и очагами демиелинизации в спинном мозге и окружающих оболочках, что сопровождается поражением и потерей аксонов. Механизм повреждения аксонов и миелина, связанный с инфекцией HTVL-1, очевидно, осуществляется через интенсивное воспаление, которое HTVL-1 вызывает в зоне поражения. Цитотоксические лимфоциты (CD8+) (CTL) обнаруживались в биоптатах, что указывает на наличие CTL-опосредованного повреждения ткани. Хотя имеется мало данных о различиях в нейровирулентности между штаммами HTVL-1 или HTVL-2, но есть данные, что отдельные вирусные гены способствуют развитию воспаления. Из исследований in vivo известно, что ген tax, кодируемый HTVL-1, трансактивирует несколько генов хозяина, в том числе, кодирующие цитокины и матриксные металлопротеиназы в лимфоцитах и астроцитах, именно эти вещества, как считается, повреждают миелин и аксоны прямо или через патологическую проницаемость гематоэнцефалического барьера [14].Лентивирусы
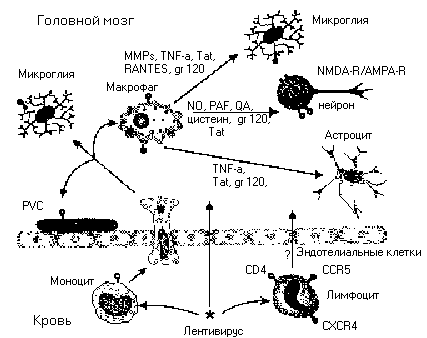
Эта группа ретровирусов привлекает повышенное внимание, ввиду эпидемии ВИЧ - СПИДа, и высокой частоты поражения нервной системы при инфицировании этими вирусами [15]. К лентивиру-сам относятся вирусы иммунодефицита, в том числе, вирус иммунодефицита человека (HIV = ВИЧ), обезьян (SIV), кошек (FIV), коров (BIV), и несколько других вирусов, поражающих копытных, например, вирус visna-maedi (VMV) [16]. Не так давно несколько исследовательских групп получили химеры SIV-HIV (SHIV), которые также проявляют нейротропность [17]. Несмотря на значительное молекулярное разнообразие каждого из нескольких генов лентивирусов, у этих вирусов много общих генетических и биологических характеристик, в том числе, общая организация ретровирус-ного генома, описанная выше. Впрочем, геномы всех лентивирусов содержат множество дополнительных генов, необходимых для способности к ин-фицированию и репликации. Лентивирусов отличает их способность реплицироваться в неделящихся клетках, например, в макрофагах [18], и нейротропность лентивирусов касается, в основном, клеток, происходящих из костного мозга, таких как микроглия и макрофаги, в то время как астроциты и эндотелиальные клетки, очевидно, инфицируются в гораздо меньшей степени [19, 20]. Широко распространено предположение, что лентивирусы проникают в ЦНС с помощью инфицированных макрофагов, пересекающих гемато-энцефалический барьер, с последующим инфицированием клеток микроглии [21] (рис. 2).
Рис. 2. Типы клеток, участвующих в патогенезе лентивирусной инфекции.
Вначале происходит инфицирование лимфоцитов и моноцитов крови, затем происходит переход инфицированных клеток через гематоэнцефалический барьер. Считается, что главный тип инфицированных клеток, проникающих в нервную систему - это моноциты/макрофаги, от которых инфекция переходит на периваскулярные макрофаги (PVC), микроглию и астроциты. Инфицированные и активированные макрофаги и микроглия продуцируют молекулы воспаления, в том числе, цитокины, хемокины и матриксные металлопротеиназы (ММР), которые способствуют гибели клетки. Кроме того, функция нейрона нарушается веществами, способными действовать как нейротоксины, к этим веществам относятся: хиноли-новая кислота (QA), цистеин, фактор активации тромбоцитов (PAF) и оксид азота (N0). Вирусные белки могут прямо или косвенно вызывать гибель нейронов, в их числе - белки Tat и env-кодируемые gp120 и gp41. У инфицированных или активированных астроцитов обнаруживается сниженный захват глутамата, таким образом повышается вероятность гибели нейрона через внешние цитотоксические механизмы. Хемокиновые рецепторы служат как ко-рецепторы при инфици-ровании лимфоцитов и макрофагов/'микроглии и могут также модулировать выживание и функционирование нейронов. Сокращения: AMPA-R - АМРА-рецептор; NMDA-R - NMDA-рецептор; TNF-a - фактор некроза опухолей а.Для некоторых лентивирусов, таких как HIV, SIV и FIV известны рецепторы к вирусам на поверхности клеток, - как в нервной системе, так и вне её, например, рецепторы CD4 или определенные хемокиновые рецепторы, либо сочетания этих рецепторов, такие как CXCR4 и CCR5 [22].
Клиника -нейробихейвориалъные и нейроструктурные особенности
Нейроструктурные и клинико-нейробихейвориальные проявления лентивирусной инфекции широко варьируют, в зависимости от конкретного вида лентивируса и факторов хозяина, например, от возраста и статуса общего иммунитета. Для всех лентивирусов характерно поражение головного мозга, приводящее к деменции или к энцефалопа-тии. Осевой причиной клинико-бихейвориальных симптомов, сопровождающих лентивирусные инфекции, считается повреждение и смерть нейронов вместе с фоновой воспалительной реакцией [20, 23]. Особенно чувствительными к повреждениям, индуцированным лентивирусами, являются отдельные популяции нейронов в базальных ганглиях и в лобной коре, именно в этих зонах макрофа-гальная инфильтрация и степень инфицированности достигают наибольшей величины [24,25]. К морфологическим изменениям относятся: потеря нейронов, вакуолизация дендритов, повреждение аксонов с одновременным глиозом, макрофагальная инфильтрация, наличие гигантских многоядерных клеток и структурно-функциональные нарушения гематоэнцефалического барьера [26, 28]. Однако, как при HIV, так и при лентивирусных инфекциях у животных часто отмечается несоответствие по тяжести между нейроструктурными нарушениями (энцефалит) и клинико-бихейвориальными проявлениями (энфефалопатия) [29, 30].Вирусные детерминанты вирулентности лентивирусов
Повреждение и гибель нейронов, очевидно, происходят за счет инфицирования их лентивирусами и за счет активации окружающих макрофагов и глии, что вызывает высвобождение веществ с потенциально нейротоксическими свойствами (таблица 2) [31]. Причинно-следственная связь между экспрессией генома лентивирусов и нейровирулентностью доказано разнообразными свидетельствами. К ним относятся сообщения о штамм-ассоциированной нейровирулентности в моделях SIV [32], VMV [33] и FIV [34] и исследования по нейротоксичности вирусных белков, выделенных при HIV, SIV, FIV и VMV [35]. Например, показано, что особая последовательность мутаций в гене env прямо влияет на развитие нейровирулетности, связанной с SIV и FIV.Таблица 2. Нейромодуляторные и нейротоксичные молекулы, участвующие в патогенезе ретрови-русного поражения нервной системы.
Источник молекул Молекулы Вирусные белки Белки, кодируемые Env (gp 120, gp 41), Tat.Nef Молекулы, обра-
зуемые в микро- фагах/микроглииТоксические факторы с низким молекулярным весом: хинолиновая кислота, глутамат, цистеин, оксид азота, супер-анион, пероксинитрит, Ntox Производные арахидоновой кислоты (простагландины Е2 и F2; лейко-триены В4 и D4; тромбоксан В2; факторы активации тромбоцитов)
Цитокины: TNF-a, ILlb, IFN-a
Хемокины: МСР-1, RANTES, MIP-la
Матриксные металлопротеиназы: ММР-2, -7, -9
Сокращения: IFN - интерферон; IL - интерлейкин; МСР - белок - хемоаттрактант макрофагов; MIP - белок, ин-гибирующий макрофаги; TNF - фактор некроза опухолей.
Для модели SIV в участке gp41, кодирующем трансмембранный белок, известны конкретные последовательности, без которых нейровирулентность не проявляется, а в модели FIV поверхностная функциональная единица gp95 представляется главным фактором, ответственным за нейровирулентность.
Указанные недавние исследования находят свое зеркальное подтверждение в сообщениях, согласно которым мутации последовательностей HIV-env, обнаруживаемые в головном мозге и в крови, коррелируют с HIV-зависимой деменцией (HIV-dependent dementia - HAD) [36,37]. Функциональные исследования показывают, что, если инфицировать макрофаги рекомбинантными вирусами, которые содержат последовательности env, полученными из головного мозга у больных HAD, то эти макрофаги начнут выделять in vitro нейротоксиче-ские молекулы[38], очевидно, посредством механизма индуцирования пути сигнального переносчика и активатора транскрипции l-Janus-киназы 1 (STAT1-JAK1) и индуцирования металлопротеи-наз [39]. Было показано, что участки протеинов, кодируемых лентивирусным геном env, прямо и косвенно проявляют нейротоксические свойства in vitro и in vivo [40, 41]. К этим последовательностям относятся участок связывания с CD-4 [42] и гипервариантные участки [43] в составе HIV-1 gp!20, причем, их действие, возможно, реализуется через механизм внешней цитотоксичности, в котором участвуют рецепторы к глутамату и потенциал-зависимая активация кальциевых (Са2+) канальцев.
Если в астроцитах трансгенных мышей имеет место экспрессия gp120, то у них отмечаются нейропатологические изменения, включающие астроглиоз, потерю нейронов, и вакуолизацию дендритов, что напоминает энцефалит при ВИЧ [44]. Для других лентивирусных белков, например, Tat при HIV и VMV, gp41 при HIV, Nef при HIV, показаны нейротоксические свойства in vitro и свойство индуцировать экспрессию генов хозяина [35]. Белок Tat находился в центре внимания многих исследований по нейропатогенезу, ввиду удивительного разнообразия его влияния на вирусную транскрипцию, на активацию генов хозяина, а также ввиду его способности вызвать смерть нейронов [45].
Число копий лентивируса или "вирусная нагрузка" является показателем с сильной прогностической силой в отношении прогрессии иммунологической болезни [46, 47], но остается невыясненной корреляция между HAD или наличием лентивирусной энцефалопатии у животных и вирусной нагрузкой на головной мозг [34, 48], хотя в зонах поражения при энцефалите часто выявляются лентивирусные антигены [20,49, 50], а вирусная нагрузка в спинномозговой жидкости имеет корреляцию с нейрови-рулентностью лентивирусов [51, 52]. Как бы то ни было, при исследовании лентивирусов in vivo, получена ценная информация о том, что системная иммуносупрессия также является важным фактором, предсказывающим прогрессирование неврологических нарушений [34, 53, 54]. В этом свете, следует провести взвешенную интерпретацию результатов исследований in vitro.
Факторы хозяина, детерминирующие нейровирулентностъ лентивирусов
Было показано, что многие вещества, выделяемые макрофагами и микроглией, могут принимать участие в патогенезе летивирусной инфекции (таблица 2). Хотя известно, что нейровирулентность лентивирусов осуществляется через активирование молекул воспаления, но до сих пор не рассматривалось влияние мутаций или полиморфизма конкретных про-воспалителъных генов хозяина. Впрочем, HIV-инфицированные лица, гетерозиготные по 32 bр делеции (b-хемокинового рецептора (CCR5), имеют меньший риск развития HIV-обусловленной деменции, по сравнению с лицами со СПИДом, не имеющими подобного полиморфизма [55]. Данный рецептор (CCR5) может быть ключевым компонентом развития лентивирусной нейровирулент-ности, ввиду того, что он необходим для инфицирования микроглии и макрофагов головного мозга вирусами HIV и SIV, а также ввиду способности CCR5 модулировать внутриклеточные сигналы при многих про-воспалительных реакциях, таких как путь STAT-JAK [56] (рис. 2). Большинство клеток в головном мозге способны вырабатывать цито-кины, но главным их источником являются активированные клетки глии, например, макрофаги, микроглия и астроциты [57]. Фактор а некроза опухолей является воспалительным цитокином, которому придается большое значение, ввиду его повышенного содержания в ткани головного мозга при инфицированности HIV, FIV и SIV и ввиду его способности влиять на высвобождение других молекул воспаления и на концентрацию внеклеточного токсического нейротрансмиттера [58]. По мере совершенствования знаний о хемокиновых рецепторах как о ко-рецепторах для лентивирусной инфекции, углубляется интерес к патофизиологическому действию хемокинов в нервной системе [59]. Исследования показывают, что белок lot, ингиби-рующий макрофаги (MIPloc) и RANTES (регулируемый при активации, экспрессируемый и секре-тируемый нормальными Т-клетками) (таблица 2) способны блокировать смерть нейронов, индуцируемую gp!20, могут влиять на Са2+ сигналы в нейронах, ингибировать лентивирусную инфекцию, а также привлекают к себе инфицированные и/или активированные макрофаги. Эти про-воспалитель-ные молекулы являются элементами внутри- и внеклеточного каскада, в котором происходит взаимодействие между клетками моноцитарно-макрофа-гальной линии и астроцитами, что способствует повреждению нейронов.
Глоссарий
Адаптационный иммунитет: иммунная реакция хозяина, осуществляющаяся посредством клеток В и Т, например, действие антител или цитоагрессивных (CD8+) Т-лимфоцитов.
Энцефалит: морфологические изменения в головном мозге, связанные с воспалением и инфекцией, в том числе, лейкоцитарная инфильтрация и активация гигантских клеток; часто отмечается присутствие вирусного генома или антигенов.
Энцефалопатия: болезнь головного мозга, которая приводит к поведенческим, когнитивным или моторным нарушениям.
env: ретровирусный ген, кодирующий оболочечные белки, например, поверхностную или трансмембранную единицы структуры.
gag: ретровирусный ген, кодирующий белки капсида и нуклеокапсида
Врожденный иммунитет: иммунная реакция хозяина, осуществляющаяся макрофагами, микроглией, дендритными клетками, тучными клетками и клетками с естественными свойствами киллеров.
Лентивирус: сложно устроенный ретровирус, для которого характерна способность инфицировать неделящиеся клетки. К лентивирусам относятся HIV, SIV, FIV.
Длинный концевой повтор (LTR): нетранслируемый участок ретровируса, который с боков ограничивает структурные гены и содержит элементы, регулирующие транскрипцию.
Миелопатия: Болезнь спинного мозга, которая приводит к моторной и сенсорной дисфункции. Нейротропизм: инфицирование вирусом нервной системы.
Нейровирулентность: болезнь нервной системы вызвана вирусом.
Открытый фрагмент для считывания: часть гена, кодирующая белок.
pol: ретровирусный ген, кодирующий обратную транскриптазу (ДНК-полимераза), интегразу и протеазные ферменты. В некоторых случаях протеаза может кодироваться геном gag.
Провирус: Промежуточная форма ретровируса в виде ДНК, обычно интегрированной в ДНК хозяина. Ретровирус: РНК-вирус (в собственном смысле этого термина), способный к обратной транскрипции.
Ретровирус типа С: простой ретровирус, часто связанный с онкогенезом, например, - MuLV, HTLV-1, HTLV-2
Механизмы гибели нейронов при лентивирусных инфекциях
Все большее внимание привлекают межклеточные процессы, посредством которых нейроны повреждаются или уничтожаются при лентивирусноых инфекциях [60]. В некоторых исследованиях показано, что гибель нейронов носит характер апоптоза, но сейчас еще неясно, играет ли ведущую роль в гибели нейронов запрограммированная смерть клетки [61]. Исследования in vitro показывают, что протеин gp!20, кодируемый геном env при HIV, может вызвать апоптоз фетальных нейронов человека через активацию процессов с участием JNK в нейронах и внеклеточно-регулируемой киназы [62]. В других сообщениях говорится, что gp!20, выделенный их штамма HIV, тропного к Т-клеткам, индуцирует апоптоз нейронов при участии р38 митоген-активируемой протеин-киназы [63]. Исследование in vitro с применением белка Tat, синтезируемого при HIV показало, что киназа Зр гликоген-синтета-зы и каспаза 9 также регулируют выживание нейрона [64]. Из этих исследований видно, что механизм и частота гибели нейронов при HIV, очевидно, определяются многими механизмами.Наметки на будущее
Ретровирусы представляют значительную проблему для здравоохранения, так как эти вирусы имеют тропность к нервной и иммунной системам. К проблемам, требующим срочного решения, относятся разрастание эпидемии ВИЧ-СПИДа и опасность ксено-инфекций от использования органов свиней и обезьян для трансплантации человеку [65]. Для разработок вирусных векторов важна устойчивость экспрессии генов, и у многих ретровирусов имеются свойства, желательные для таких разработок, а именно способность встраиваться в геном хозяина и реплицироваться в неделящихся клетках [66]. Но при этом существуют и проблемы безопасности, связанные с возможной разработкой вирулентных ретровирусных векторов [67] и с потенциальной токсичностью лентивирусных белков, таких, как Tat [68], который предлагается в качестве переносчика генов. Более того, лентивирусы, паразитирующие на животных, и их векторы способны инфицировать человеческие клетки и вызывать гибель лимфоцитов, макрофагов и нейронов [69]. Тем не менее, с рекомбинантными нереплицирующимися ретровирусными векторами, способными инфицировать лишь определенные клетки, связаны надежды на лечение таких неврологических нарушений, как болезнь Паркинсона, инсульт и опухоли головного мозга.К разряду развивающихся ретровирусных инфекций можно отнести эндогенные ретровирусы [70] и вирусы spuma [3]. Хотя в природных условиях вирус spuma редко вызывает болезнь у своих хозяев, но он способен вызывать нейродегенерацию, будучи экспрессированным как трансгены у мышей, и может инфицировать разные виды, в том числе людей, поэтому применение его как возможного вектора вызывает сомнения [71]. Эндогенные ретровирусы (EVR) - это интересная группа ретровирусов, обнаруживаемая у большинства видов млекопитающих, в том числе - у людей, мышей, приматов и кошек, и эти вирусы могут составлять до 1 % генома хозяина [72]. У EVR обычно дефектная репликация, но некоторые могут рекомбинировать со своими экзогенными гомологами, с образованием вирусов, способных к репликации, что показано на примере FeLV и MuLV. Обсуждается участие EVR в болезнях человека, в том числе, недавно ECR связали со множественным склерозом и другими аутоиммунными заболеваниями с неврологическими проявлениями [73].
Нейродегенерация в результате хронического воспаления является кардинальной особенностью нейротропных ретровирусных инфекций. Многие ретровирусы сейчас выделены как отдельные молекулярные инфекционные клоны, и поэтому имеется теоретическая возможность прерывать in vivo конкретные сигнальные механизмы, которые запускаются ретровирусами и кончаются нейродегенерацией. Провоспалительные и нейротоксические молекулы хозяина принимают участие в каскаде взаимодействий, имеющих результатом нейровирулентность ретровирусов. Считается, что эти молекулы принимают участие в развитии таких видов нейродегенеративной патологии человека, как рассеянный склероз, инсульт, болезнь Альцгеймера, травмы головного мозга [74]. Следовательно, механизмы, посредством которых ретровирусы индуцируют воспаление и повреждение тканей, могут подсказать идеи о патогенезе других заболеваний головного мозга, сопровождающихся воспалением.
Полная библиография к статье находится в редакции журнала
Апрель 2002 г.