Опубликовано в журнале:
АКТИВНЫЕ ФОРМЫ КИСЛОРОДА В РЕАКЦИЯХ ПОВРЕЖДЕНИЯ БИОМОЛЕКУЛ
Молекулярный кислород сам по себе обычно не вступает в неконтролируемые химические реакции внутри организма, для его активации нужны ферментативные процессы – главные ферменты метаболизма кислорода у млекопитающих: оксидазы и оксигеназы. Но в каталитических центрах этих ферментов кислород испытывает превращения до конечных соединений, не выделяясь в среду и не подвергая опасности органические макромолекулы клетки, повреждающими же агентами являются активные формы кислорода (АФК), образующиеся в ряде физико-химических процессов в организме. Главные активные формы кислорода (Осипов и др., 1990):
Синглетный кислород образуется в реакциях фотоокисления в присутствии так называемых фотосенсибилизаторов: флавины, гематопорфирин, хлорофилл и др., а также при дисмутации супероксидных радикалов (Khan,1970). Синглетный кислород агрессивен в отношении биосубстратов, в особенности в отношении молекул с двойной связью; конечным итогом таких реакций обычно является образование гидроперекисей органических молекул – один из важнейших в процессах перекисного окисления ненасыщенных липидов в биомембранах (Осипов,1990, Roschupkin, Pelenitsin, Potapenko etc, 1975).
В присутствии металлов переменной валентности эти продукты запускают цепные реакции окислительной деградации биомолекул (Vladimirov etc., 1980). Главным защитным механизмом такого процесса является бета-каротин, переводящий синглетный кислород в триплетный, однако, обычная вода и токоферол также способны вызывать такой процесс инактивации синглетного кислорода (Duran, 1982, Mercel, Kearns, 1972) . В клинической практике синглетный кислород участвует в кожных проявлениях некоторых генетических заболеваний – порфирий (Frank, 1982), а также в процессах эритемы при ультрафиолетовом облучении кожи при приеме лекарств, обладающих фотосенсибилизирующим действием.
Во всех аэробных клетках в процессе присоединения одного электрона к молекуле кислорода образуются супероксидный анион-радикал - О2- и его протонированная форма – гидроперекисный радикал - НО2*; оба они порождают ряд других активных форм кислорода. Образование этих АФК наиболее существенно вблизи цепей переноса электронов – дыхательная цепь, микросомы и, в растительных клетках, хлоропласты (Каган, Сербинова, Минин и др, 1985, Rosen, Finkelstein, Rauckman, 1982).
Эти АФК играют также важную роль в защитных - неспецифических иммунных механизмах организма: они выделяются в ходе активации при инфекционных и иных воспалительных процессов фагоцитирующих клеток (нейтрофилы, макрофаги, моноциты, эозинофилы) (Маянский, Маянский, 1983, Edwards,. Hallett, L loyd etc, 1983, Klebanoff, Clark, 1978). Супероксидный радикал может прямо инактивировать адреналин, аскорбиновую кислоты (Bhuyan, Bhuyan, 1977), а более активный гидроперексиный радикал – реагирует с линолевой, линоленовой, арахидоновой кислотами, окисляя их до гидроперекисей (Bielsky, Arudi, Sutherland, 1983). Образованию гидроперекисного радикала способствует закисление среды, он также свободно проникает через биомембраны, так как не несет заряда.
Гидроперекиси липидов являеются весьма активными соединениями и обладают высокой биологической агрессивностью. Для протекания цепного окисления липидов в биологических мембранах совершенно необходимы переходные металлы, в частности, ионы железа (Ванин,1967, Владимиров, Арчаков, 1972). Простым и доступным методом определения продуктов перекисного окисления липидов является реакция с тиобарбитуровой кислотой (Marcuse, Johnston, 1973). Главным механизмом защиты организма от данных форм АФК является фермент супероксиддисмутаза (СОД), активность ее обычно достаточна, чтобы инактивировать их в месте образования, не допуская диффузии в среде макромолекул ткани.
Для определения концентрации супероксидных анион-радикалов используют методы электропарамагнитного резонанса (ЭПР-резонанс) и реакции оксиления интенсивно поглощающих соединений: цитохрома С, нитросинего тетразолия, адреналина и др. (Massey, 1959, Beyer, 1987, Misra, Frodowich, 1972).
Для доказательства специфичности этих реакций используют их подавление супероксиддисмутазой.
Дисмутация супероксидных анион-радикалов под действием СОД в биологических тканях ведет к образованию перекиси водорода, способной легко проникать через мембраны клеток. Перекись водорода обнаруживается при фагоцитозе, при работе митохондрий и микросом (Rosen, Finkelstein, Rauckman, 1982, E dwards, Hallett, Lloyd etc, 1983). В присутствии ионов переходных металлов (например Fe2+) перекись водорода может давать высоко активный гидроксильный радикал (* ОН). Этому процессу препятствуют главные высоко активные ферменты антиоксидантной защиты организма: каталаза и глутатион-пероксидаза. Измерение перекиси водорода в биосубстратах проводят обычно методом титрования перманганатом калия (Bonnichsen, 1948), реакцией с молибдатом аммония (Королюк, Иванова, Майорова, Токорева, 1988) или прямой спектрофотометрией при длине волны равной 240 нм (Bergmeyer, 1955, Cohen, Dembiec, Marcus, 1970).
Используют также пероксидазную реакцию, в ходе которой изменяется окраска индикатора, например, индигокармина (Frew, Jones P, Sholes, 1983), и флюориметричсекие методы (Perschke, Broda, 1961, Keston, Brandt, 1965).
Высокой реакционной способностью обладает гидроксильный радикал (* ОН), образующийся из перекиси водорода в присутствии ионов переходных металлов. Высокая реакционная способность определяет преимущественно местное воздействие этой форм АФК. Прямое повреждение ДНК при этом характеризуется разрывом цепи (180); с другими биомолекулами * ОН образует вторичные свободные радикалы, в том числе перексиные соединения липидов (Азизова и др, 1985, O`Connel, Garner, 1983, Rowley, Halliwell, 1982).
Главные типы повреждений биомолекул * ОН: отрыв атома водорода (таким образом повреждается лецитин – главный компонент биологических мембран, а также сахара в составе нуклеозидов ДНК); присоединение к молекулам по двойным связям (взаимодействие с пуринами и пиримидинами ДНК и РНК, в том числе с образованием вторичных радикалов); перенос электронов также является патогенным механизмом действия * ОН (Anbar, Neta 1967). В образовании гидроксильного радикала важное значение имеют ионы металлов с переменной валентностью, в первую очередь ионы железа (Flitter, Rawley, Haliwell, 1983, Floyd, 1983, Jacobs, 1977).
Ионы железа входят в большом количестве в состав организма (гемоглобин, миоглобин и пр.); в крови они находятся в связанной форме с трансферрином.
Снижение количества железо-переносщих белков и повышение свободного железа крови может вести к стимулированию формирования свободных радикалов; своевременная диагностика и профилактика такого состояния является важным моментом программ диагностики и профилактики старения, в частности, американской ассоциации Life Extension Foundation.
Отмечено, однако, формирование гидроксильного радикала и под действием связанного железа – лактоферрина (Ambruso, Johnston, 1981, Bannister, Bannister, Hill, Thornalley, 1982), а также при действии гемоглобина на перекись водорода (Banatti, Morelli, Guida, de Flora, 1983). Использование хелатосвязывающих агентов дает лечебный эффект при состояниях, при которых предполагается участие гидроксильного радикала в патогенезе заболевания: связывающий ионы железа десферриоксиамин эффективен при воспалительных процессах в легких (Ward, Till, Kunkel etc.,1983) и при аутоиммунных процессах (Bowern, Ramshow, Clark etc., 1984, Clark, Hunt, 1983).
АФК могут образовываться также и при многих иных процессах в организме. Так, например, формирование АФК обычно сопровождает процесс инактивирования в организме ксенобиотиков (Mustafa, 1990, Sinha, Mimnough, 1990), потенцируя их повреждающий эффект.
Главные методы определения * ОН: ЭПР, хроматография биомолекул для определения результатов их изменения под действием * ОН и прямые химические методы определения таких агентов, важным является также метод хемолюминесценции биомолекул, обычно с использованием активаторов хемилюминесценции, наиболее известными из которых являются люминол, люциноген (Allen, 1982).
ЗАЩИТА ОРГАНИЗМА ОТ АФК
СУПЕРОКСИДДИСМУТАЗА (СОД). Супероксиддисмутаза является важнейшим элементом антиоксидантной защиты организма. Это фермент из двух субъединиц с общей молекулярной массой 32 кДа, содержащий по одному атому меди и цинка (существует также марганец-содержащая СОД, обнаруженная в печени крысы и человека; в бактериальных клетках обнаружена железо-содержащая СОД). Фермент ускоряет распад О2- на 4 порядка. Активность СОД обычно определяют по ингибированию содержащих СОД биологическими экстрактами и жидкостями образования под действием О2- окрашенных продуктов из субстратов, например, тетразолия нитросинего (Чевари., Чаба., Секей 1981).
Источником О2- обычно является система феназинметасульфат + НАД*Н или ксантин + ксантиноксидаза. За единицу активности СОД принято ее количество, тормозящее в 2 раза восстановление цитохрома с при реакции ксантиноксидазы (0,003 ед/мл), ксантина (2 мкмоль/мл) и цитохрома с - 0,6 мкмоль/мл (McCord, Fridovich, 1969).
Вторым эшелоном защиты организма от АФК являются пероксидаза и каталаза.
КАТАЛАЗА. Каталаза расщепляет перекись водорода, до которой дисмутирует супероксидный радикал, до молекул воды и молекулярного кислорода. В клетках каталаза в основном сосредоточена в пероксисомах, в которых содержатся и ферменты, продуцирующие перекись водорода, необходимую в ходе ряда процессов жизнедеятельности организма, в частности, в процессах неспецифической иммунной защиты.
ПЕРОКСИДАЗА. Пероксидаза, в особенности глутатион-пероксидаза, широко распространена в клетках животных и растений. Глутатион-пероксидаза состоит из 4 субъединиц в каждой из которых содержится по молекуле селена. В клектах этот фермент располагается в цитозоле и матриксе митохондрий.
Активность глутатион-пероксидазы зависит от содержания глутатиона клетки, что, в свою очередь, определяется активностью глутатионредуктазы и концентрацией НАДФ*Н, который образуется в пентозофосфатном метаболическом цикле. Лимитирующими органами по активности каталазы, являются легкие, мышцы, глаза.
ДРУГИЕ ФОРМЫ ЗАЩИТЫ ОТ АФК
В защите от АФК в организме участвуют и многие другие молекулы и ферментные системы (Осипов, Азизова, Владимиров, 1990).
Классические в настоящее время антиоксиданты – витамин Е, витамин А и каротиноиды, активны почти ко всем АФК, но их вклад в общую антиоксидантную активность организма не слишком велик.
Из других жирорастворимых агентов антиоксидантной активностью обладают стероидные гормоны, билирубин; из водорастворимых – церрулоплазмин (влияя на свободное железо крови), трансферрин, альбумин, SH-группы белков.
Аскорбиновая кислота инактивирует свободные радикалы, образуя неактивный радикал (семидегидроаскорбат), она же является кофактором пероксидазы, (фермент аскорбат-пероксидаза).
Глутатион, присутствуя в клетках в высоких концентрациях, также является акцептором гидроксильного иона и синглетного кислорода, кроме того, он же является кофактором глутатион-пероксидазы и глутатион-редуктазы. Мочевая кислота присутствует в крови в достаточных количествах, чтобы эффективно акцептировать синглетный кислород и гидрок-сильный радикал. Аналогичными эффектами обладают этанол, маннит, глюкоза и некоторые другие органические молекулы.
АФК В НЕСПЕЦИФИЧЕСКОМ ИММУНИТЕТЕ И ВОСПАЛЕНИИ
Формирование АФК – важный защитный механизм, лежащий в основе неспецифического иммунитета: фагоцитоз приводит к многократному увеличению содержания АФК в фагоцитирующих клетках с одновременным повышением потребления кислорода в 20 и более раз ("дыхательный взрыв" (Маянский, Маянский, 1983, Klebanoff, Clark, 1978).
Участие АФК в процессах фагоцитоза достаточно сложное.
Фагоцитирующая клетка активируется бактериальными клетками (или механическими частицами, лектинами и пр.), что сопровождается активацией фермента плазматической мембраны – НАДФ*Н-оксидазы с формированием из свободного молекулярного кислорода О2- (Babior, 1978, Badway, Karnovsky, 1980). В процессе генерации АФК участвуют ФАД-содержащий флавопротеин и цитохром b .
В конечном счете с участием ионов железа происходит дисмутация АФК до перекиси водорода.
Кроме того, миелопероксидаза нейтрофилов приводит к образованию гипохлорита, хотя последний не является главным в антибактериальной защите организма.
Высвобождние АФК в ходе "дыхательного взрыва" происходит как в фагосомы, так и в среду, что инактивирует как бактериальные клетки, так и может повреждать сами фагоциты, а также нормальные ткани.
Для защиты от АФК нейтрофилы содержат каталазу и глутатион-пероксидазу (Klebanoff, Clark, 1978).
Активация нейтрофилов сопровождается также при любых явлениях некроза ткани, в том числе микроинфарктах (Клебанов и др., 1984, 1987 ).
Участие гидроксильных радикалов подтверждено в патогенезе ревматоидного артрита, при этом фагоциты активируются иммунными комплексами в синовиальной жидкости (Bennett, Eddie-Quartey, Holt, 1973, Bennett, Skosey, 1977), при этом введение СОД в полость сустава оказывается терапевтически высоко эффективным.
Окисленные липиды обладают антигенными свойствами, запуская аутоиммунные процессы повреждения тканей (Деев и др.,1987, Hejnecke, 1987).
Бронхоконстриктивные заболевания легких, обычно сопровождающиеся хроническими воспалительными процессами, являются второй важнейшей патологией, в которой участие АФК достаточно важно, как это представляется на настоящий момент.
АФК сами способны вызывать бронхоконстрикцию, кроме того, гистамин в ходе развития хронических обструктивных заболеваний легких способен вызывать продукцию АФК вследствие извращения реакции на него нейтрофилов – при бронхиальной астме растормаживается ингибирующее действие гистамина на нейтрофилы; сходным образом действует и ацетилхолин (Коган, 1999). Увеличивается также генерация АФК в ходе приступа бронхиальной астмы, причем обнаруживается параллелизм в тяжести астматических приступов и генерации АФК нейтрофилами и повышением содержания в крови ПОЛ. Имеет место также лечебный эффект антиоксидантной терапии при этом заболевании.
Известен механизм усиления продукции лейкоцитами АФК при обострении бронхиальной астмы (Коган, 1999): он заключается в снижении АФК-ингибирующей функции тромбоцитов в отношении лейкоцитов и стимуляции лейкоцитов под действием ацетилхолина, а также в извращении тормозащего действия гистамина на лейкоциты.
Важное значение имеет утрата лейкоцитами особого недавно открытого отечественными исследователями эффекта – супероксидингибирующего действия углекислого газа (Коган, 1999).
Так, было показано, что влияние углекислоты на многие типы тканей ведет к снижению в 2-4 раза продукции ими генерации супероксидных радикалов.
Этот эффект может лежать в основе тренирующего и лечебного действия гиперкапнических методов терапии и профилактики бронхиальной астмы. В то же время, в ряде случаев такой лечебный эффект отсутствует. Было показано, что в большинстве таких случаев отсутствует и эффект углекислого газа на подавление продукции супероксидных радикалов лейкоцитами таких больных.
Выше обсуждались также эффекты участия АФК в хроническом воспалении, всегда присутствующем при бронхоконстриктивных заболеваниях легких.
Таким образом, воспалительные и констриктивные легочные заболевания – еще один важный патологический процесс, в котором явно принимают участие АФК.
АФК В СЕРДЕЧНО-СОСУДИСТОЙ ПАТОЛОГИИ
Участие АФК в сердечно-сосудистой патологии в настоящее время не оставляет сомнений. Показано усиление процессов перикисного окисления липидов (ПОЛ) в ишемизированном миокарде (Коган, Кудрин, Лосев, 1987, Kogan, Kudrin, 1980). Между продукцией в тканях миокарда перекиси водорода, повреждающем действием перекиси и повышением чувствительности к ней ишемизированных тканей устанавливается порочный круг; повреждению способствует также эмиграция лейкоцитов в зону воспаления и снижение в ней активности ферментов антиоксидантной защиты тканей (Коган, 1999).
Ишемия, парадоксально, не отражается на продукции АФК, в то же время, выраженно повреждая аэробные ткани вследствие недостатка обычного кислорода. Так, показано, что снижение кислорода в 100 раз по отношению к атмосферному, снижает продукцию АФК макрофагами не более чем на 25% (34ж). В то же время, активность СОД, глутатионпероксидазы в ишемизированной области снижается уже в первые минуты ишемии и сохраняется в течение всего периода эксперимента по ишемии миокарда (Лапкин и др., 1982).
Активация селеном активности глутатионпероксидазы обладает протективным эффектом на экспериментальный инфаркт (Коган, 1992), снижая зону морфологически видимого инфаркта и изменения ЭКГ.
В противоположность, активация продукции АФК форболмиристат ацетатом вызывает резкую дисфункцию сердца: уменьшение в 6 раз сердечного индекса, ЭКГ-регистрируемую ишемию миокарда и гибель животных (Rao, Mueller, 1983).
Все эти эффекты могут быть ревертированы внутривенным введением СОД и каталазы или же снижением количества нейтрофилов (антинейтрофильными антителами) как основного источника в норме АФК в организме. Введение СОД снижало также размеры инфаркта миокарда после коронароакклюзии у крыс (Коновалова, Лапкин, Бескровнова, 1989).
СОД защищает, видимо, как от первичных повреждений ишемизированной ткани в ходе начавшейся коронароакклюзии, так и от усугубления повреждения после восстановления кро-вотока - когда повышение кислорода в ткани при восстановлении кровотока ведет к активации продукции АФК и усилению повреждения.
Имеются данные об активации СОД – повышении активности СОД в оттекающей от зоны инфаркта крови, при этом повышается и содержание перекиси (Rao, Mueller, 1983).
Косвенным показателем участия продуцируемых лейкоцитами АФК в патогенезе инфаркта служит и известный факт ухудшения прогноза при повышении степени лейкоцитоза при инфаркте; генерация АФК нейтрофилами и тяжесть стенокардии также параллельны.
Включение коэнзима Q (антиоксиданта, работающего на митохондриальном уровне), улучшает лечебные эффекты при стенокардии.
Интересные данные о влиянии коэнзима Q на эффекты гипербарической оксигенации: без антиоксидантов гипербарическая оксигенация приводит к временному ухудшению и стимулированию приступов стенокардии, с одновременным увеличением продукции АФК нейтрофилами; антиоксидантная терапия снимает эти эффекты, сохраняя лечебное действие гипербарической оксигенации (Аль-Хадиди, 1987).
Антиоксидантная терапия эффективна также в профилактике стенокардитических приступов и достоверно повышает толерантность к физической нагрузке (велоэргометрии).
Все выше перечисленные данные позволяют говорить о кислородном свободнорадикальном механизме аутоагрессии при ишемической болезни сердца и о целесообразности включения в лечение этих заболеваний средств, регулирующих продукцию и инактивацию АФК.
АФК В ПРОЦЕССАХ КАНЦЕРОГЕНЕЗА
Вопрос участия АФК в процессах возникновения опухолей постоянно привлекал к себе внимание исследователей, однако, до настоящего времени он фактически не однозначно не решен. Конкретный механизм индукции опухолей свободными радикалами мало понятен. Предполагают, что АФК повреждают хроматин, ДНК, мебраны, изменяют регуляцию внутриклеточного кальция и пр. (Кольтовер, 1998, Пескин, 1997, Dogru-Abbasoglu, Tanger-Toptani, Ugernal etc, 1997, Papa, Skulachev, 1997, Yan, Levine, Sohal, 1 997). Важным также является разнонаправленность изменений антиоксидант-ного статуса в различных органах (Кольтовер, 1998, Dogru-Abbasoglu etc., 1997, Papa, Skulachev, 1997), что соответствует и различной чувствительности к химическим канцерогенам и ионизирующему облучению.
С возрастом изменяется скорость накопления соматических мутация в разных тканях, причем она выше в печени, чем в мозгу (Dolle etc., 1997). Все это позволяет ряду авторов говорить о возможности использования антиоксидантов в качестве геропротекторов и средств коррекции возрастной патологии, в том числе опухолей, так как риск их возникновения выраженно растет с возрастом (Anisimov, Mylnikov, Khavinson, 1998, Cutler, 1991, Harman, 1994, Shigenaga, Hogen, Ames, 1994).
В настоящее время изучается и другой аспект участия АФК в процессах канцерогенеза, а именно – контроль АФК естественных цитотоксических реакций (Чекнеев, 1999).
Известно, что АФК участвуют в деструкции клеток-мишеней в процессах их взаимодействия с ЕК - естественными киллерами (Cameron, 1986, Saito etc., 1987). Антиоксиданты – диметилсульфоксид, диметилмочевина, этиловый спирт, этиленгликоль, снижают активность ЕК (Suthanthiran etc., 1984), как и обработка мононуклеаров миелопероксидазой (El-Hag, Clark, 1984) а ингибирование активности СОД хелатирующими комплексами усиливает цитотоксическую реакцию (Van Kessel etc., 1987).
В то же время, перекись водорода регуляторно повышает активность ЕК и выработку интерферона (Munakata etc., 1985).
В целом, видимо, имеет место сложная динамическая регуляция иммунной системы с участием АФК, в которой принимают участие, кроме выше описанных, также Р-белки сыворотки крови, обладающие СОД-активностью (Кульберг и др., 1986, Петяев, Кульберг, 1988) и комплексированные с ними сывороточные гамма-глобулины (Кульберг, Петяев, Замотаева, 1988), снижающие антенсивность свободнорадикальных процессов. В экологическом плане рассматривают также участие глобальной экспансии цианобактерий, оказывающих негативное влияние на активность ЕК-иммунитета (Кульберг, 1994, Чекнев, Кульберг, 1995).
УЧАСТИЕ АФК В ПРОЦЕССАХ СТАРЕНИЯ ОРГАНИЗМА
Старение организма – сложный и многогранный процесс, он не может быть в принципе сведен к одному конкретному механизму (наши ссылки), но среди групп влияний, играющих важную роль в старении, безусловно важное значение имеет механизм повреждения биомолекул самыми разнообразными внешними влияниями.
Среди таких влияний многие авторы давно пытались выделить наиболее значимые. Среди таких попыток один из наиболее значимых факторов оказался окислительный метаболизм – оказалось, что его интенсивность обратно пропорциональна продолжительности жизни для многих далеко отстоящих друг от друга видов, хотя некоторые исследователи отмечают, что это соблюдается достаточно хорошо только для относительного (на единицу массы) значения его, причем в расчете на единицу активности супероксиддисмутазы – ключевого фермента антиоксидантной защиты организма (Гродзинский и др., 1987, Дупленко, 1985, Azbel, 1994). Эти факты послужили основой для выдвижения свободнорадикальной теории старения.
Основное положение свободнорадикальной теории старения сформулировано было в 1954 году D.Harman, предположившим, что универсальной причиной старения служит свободнорадикальное окисление липидов, жиров и белков всех организмов кислородом воздуха (Harman, 1991). В дальнейшем было выполнено значительное число самых разнообразных работ в этой области, но каких-либо однозначно интерпретируемых результатов получено не было.
Так, митохондрии (основной потребитель кислорода клеток) старых животных не отличались значимо от таковых молодых (Кольтовер, 1996, 1998) по генерации супероксидного радикала, хотя отмечена структурная и функциональная неоднородность митохондрий старых животных, приводящая к значительным разбросам в результатах исследований и тенденция к повышению генерации АФК митохондриями старых животных все же имеет место (Кольтовер, 1996,Hagen etc.,1997).
Аналогично, микросомы – другой значимый источник супероксидных радикалов, в старости генерируют в 2 раза меньшие количества АФК, снижение же активности СОД цитоплазмы происходит всего на 40%, что явно компенсирует эти изменения (Гуськова, Виленчик, Кольтовер, 1998). В отдельных случаях активность СОД тканей может даже повышаться с возрастом (Dehaan etc., 1995). Активность ферментов антиоксидантной защиты, в принципе, зависит от нейро-эндокринной регуляции и может, таким образом, подтверждать не теорию повреждений, а регуляторную теорию старения (Bolzan etc., 1995).
Активность СОД мозга долгоживущих мышей оказывается выше, чем для короткоживущих линий (Kellog, Fridowich, 1976). Генетически измененные дрозофилы с повышенной активностью СОД живут дольше нормальных мух (Orr, Sohal, 1994). Ингибиторы свободнорадикальных механизмов (дибунол и пр.) увеличивают на 20-30% длительность жизни мышей (Обухова, Эммануэль, 1983, Harman, 1991). Для людей, однако, можно видеть противоположное – синдром Дауна с укорочением продолжительности жизни вдвое сопровождается ростом активности СОД тканей (Dehaan etc., 1995). Фармакокоррекция системы антиоксидантной защиты организма также мало достоверна – активность обычных ферментов (СОД, каталаза, перкосидаза) очень высока и специфична, поэтому способы фармакологического "тушения" антиоксидантов, видимо, мало эффективны в принципе. Более эффективны непрямые методы, например, нормализации функции митохондрий, где эффективны окись азота, кортикостероиды, тиреоидные гормоны (Koltover, 1995).
В то же время, с возрастом безусловно повышается содержание в тканях человека и животных продуктов окислительного повреждения макромолекул, в том числе ДНК (Гродзинский и др.,1987, Фролькис, Мурадян, 1988, Ozawa , 1997).
С нашей точки зрения, впрочем, это указывает лишь на снижение скорости метаболизма для организма в целом. Повреждения ДНК, однако, могут играть важную роль в мутагенезе в старости (Lee, Weindruch., Aiken, 1997, Ozawa, 1997).
Возможно, АФК стимулируют и апоптоз – программируемую гибель клеток, путем раскрытия каналов мембраны для белка, находящегося в межмембранном пространстве и запускающего этот процесс (Kroemer, 1997). АФК, несомненно, главный мутаген аэробных клеток.
АФК могут снижать скорость клеточного деления, возможно, влияя на так называемый "пределе клеточных делений Хейфлика" – культуральный феномен остановки клеточного роста после ряда пассажей культур на свежую среду.
Возможно влияние АФК на ускоренное укорочение концевых (теломерных) участков ДНК – процесс, которому придают важную роль как в клеточном делении, так и в его ограничении (Orr, Sohal , 1994).
Интересны данные получены при параллельном изучении продукции АФК тканями молодых и старых животных (Анисимов и др., 1999). Показано, что в головном мозгу старых животных почти в 2 раза снижается активность СОД; это, однако, не приводило ни к повышению концентрации диеновых конъюгатов ни к повышению показателей перекисного окисления белков или общей антиокислительной активности (содержание аскорбиновой кислоты, токоферола, мочевой килсоты, тиоловых соединений – глутатиона и др.), концентрация же шиффовых основания даже снижалась.
В то же время, в печени старых животных существенно повышалась концентрация шиффовых оснований и продуктов перекисного окисления белков, также при снижении активности СОД в 2 более раза; но не изменялись уровень общей антиокислительной активности и содержание диеновых конъюгатов.
В сыворотке старых крыс существенно повышалось содержание продуктов перекисного окисления липидов и белков при снижении активности СОД и общей антиокислительной активности сыворотки крови.
В литературе отмечены данные о существенном снижении в сыворотке крови пожилых и старых лиц (60-97 лет) уровня глутатиона и повышение продуктов перекисного окисления липидов (Nuttal, Martin, Hutchin etc, 1998). Возможно, накопление продуктов перекисного окисления липидов в мозгу играет роль и в патогенезе возрастной патологии мозга, включая болезнь Альцгеймера (Romero etc., 1998). Интересно, что эти изменения идут на фоне существенного снижения с возрастом генерации АФК в тканях старых животных и человека. В связи с этим снижение активности ферментов антиоксидазной защиты может в этих условиях быть результатом чисто регуляторных изменений – приспособления к пониженной продукции АФК.
Накопление же конечных продуктов повреждения тканей может быть результатом иных процессов – прежде всего, результатом снижения метаболизма и физиологической регенерации большинства самообновляющихся тканей в старости – в этих условиях снижается обновление всех макромолекул и биологических структур, что создает возможности для персистенции измененных форм макромолекул вследствие общего увеличения длительности их жизни.
Сейчас все больше склоняются к мнению, что наиболее значимым при старении оказывается усиленное перекисное окисление белков, а не липидов, чему способствует снижение активности СОД как результата снижения общей продукции АФК (Анисимов и др., 1999).
Общий анализ данных об участии АФК в процессах старения и сопровождающих старение заболеваний позволяет ряду авторов утверждать, что повреждение под действием АФК макромолекул приводит к мутациям, нестабильности генома в целом и развитию ряда возрастных патологий: рак, сердечно-сосудистые заболевания, возрастная иммунодепрессия, дисфункция мозга, катаракта и др. (Голубев, 1996, Кольтовер, 1998, Cutler, 1991, Dogru-Abbasoglu etc.,1997, Shigenag aetc., 1994).
Конкретный механизм индукции старения свободными радикалами мало понятен. Предполагают, что АФК повреждают хроматин, ДНК, мембраны, коллаген, изменяют регуляцию внутриклеточного кальция и пр. (Кольто-вер, 1998, Пескин , 1997, Dogru-Abbasoglu etc., 1997, Papa, Skulachev, 1997, Yan, Levine,etc., 1997). Важным также является разнонаправлен-ность изменений антиоксидантного статуса в различных органах (Кольтовер, 1998, Dogru-Abbasoglu etc., 1997, Papa., Skulachev, 1997), что соответствует и различной чувствительности к химическим канцерогенам и ионизирующему облучению. Скорость накопления соматических мутация в разных тканях может существенно различаться с возрастом, причем она выше в печени, чем в мозгу (Doll etc., 1997).
Все это позволяет говорить о возможности использования антиоксидантов в качестве геропротекторов и средств коррекции возрастной патологии (Anisimov etc., 1998, Cutler, 1991.
Данные, полученные на трансгенных дрозофилах с дополнительными копиями гена СОД и каталазы показали увеличение средней и максимальной продолжительности жизни (Orr, Sohal, 1994); трансгенные мыши с избыточной экспрессией СОД, каталазы и глутатион-пероксидазы обладали повышенной резистентностью к оксидативному стрессу, при снижении же активности этих генов мыши имели признаки преждевременного старения (Ho etc., 1998).
НЕКОТОРЫЕ ДРУГИЕ ЭФФЕКТЫ АФК
В последние годы все большее внимание привлекают физиологические эффекты АФК, которые оказались на удивление разнообразными.
Так, кислороду и АФК придают важное значение в процессах эволюции и видообразования (Скулачев, 1998), у целого ряда клеток АФК вызывают повышение клеточного деления (Nashio, Watanabe, 1992, Dypbukt etc., 1994, Bhunia etc., 1998, Lee S-L., Wang, Fanburg, 1998, Lee S-L., Wang W-W-, Fanburg, 1998), причем показано, что это регуляторный эффект, реализуемый через специфические белки (NF-kB, c-Jun, p21, p44MAPK, c-fos).
Достаточно изучен процесс регуляции АФК сосудистого тонуса (Rubanyi, 1998). Общеизвестен и хорошо изучен механизм "дыхательного взрыва" фагоцитов, являющийся главным компонентом неспецифической иммунной защиты организма.
Продуктам перекисного окисления липидов (ПОЛ) придают важное значение в процессах нормального обновления клеток и клеточных мембран – в поддержании структурного гомеостаза (Зенков, 1993).
Действие внешних прооксидантов, повышенное потребление кислорода, ионизирующее и ультрафиолетовое облучение, загрязнение воздуха, воды и продуктов, недостаток естественных антиоксидантов (витамины Е, К, А, селен и пр.), врожденная недостаточность ферментов антиокосидантной защиты, другие состояния могут приводить к напряжению системы антиоксидантной защиты организма и вызывать так называемый "оксидативный стресс", проявляющийся на молекулярном, клеточном и организменном уровне (Sies, 1991).
Подобный стресс, видимо, является патогенетическим моментом в развитии воспалительных, бронхо-легочных и сердечно-сосудистых заболеваний (Меньшикова, Зенков, 1991).
ЭХАС – НОВЫЙ МЕТОД ВОЗДЕЙСТВИЯ НА АНТИОКСИДАНТНУЮ СИСТЕМУ
ПОЛУЧЕНИЕ И СВОЙСТВА ЭХАС
Существует путь фармакологической стимуляции антиоксидантных механизмов защиты организма. Такие фармакологические агенты оказались высоко эффективны в эксперименте и клинике как геропротекторы, проявляющие параллельно по данному же механизму свойства антиканцерогенов, радиопротекторов и вообще биостимулирующих веществ. Сейчас становится достаточно ясно, что важнейшим механизмом действия таких средств является влияние на окислительно восстановительный потенциал воды и макромолекул. До последнего времени было затруднительно использовать практически данный путь коррекции ОВП, так как расчетные требуемые количества препарата, который должен поступать длительно в организм, оказываются достаточно велики.
Однако, в настоящее время ситуация принципиальным образом меняется в связи с разработкой новых способов получения веществ с требуемыми показателями ОВП, совершенно безвредных для организма на основе обычной воды - электрохимически активированные системы – ЭХАС (Бабенко, Гойницкий, 1976, Бах, Зубкова, 1997, Бенина, Чегунова, 1974, Бобоходжаев, 1973, Гончарик , 1973, Громашевская и др., 1973).
Есть способ, позволяющий быстро, надежно и в широких масштабах изменять ОВП и рН внутренних сред организма: использование ЭЛЕКТРОХИМИЧЕСКИ АКТИВИРОВАННЫХ СИСТЕМ (ЭХАС) в виде водных растворов или даже просто воды, которая применяется в количестве 1-1,5 кг в сутки.
Метод ЭХАС позволяет вводить в организм стандартизированную по показателям ОВП и рН воду, обогащенную за счет электро-химической обработки по ОН- или Н+ группам и с регулируемым ОВП.
Одновременно в ходе обработки вода приобретает ряд ценных свойств - обеззараживается, дегазируется от хлора, детоксицируется за счет окисления-восстановления токсических соединений и удаляются путем перевода в нерастворимый осадок соли тяжелых металлов.
Известны три основных действующих фактора для ЭХАС:
- образование устойчивых химических соединений (кислот в анолите и щелочей в католите), изменяющих рН жидкости, чему в начале и уделяли главное внимание многие исследователи;
- формирование неустойчивых (метастабильных) суперактивных соединений (высоко окисленные формы ионов и молекул, свободные радикалы), что оказывает физико-химическое и тренирующее действие на системы защиты организма и изменяет ОВП;
- возникновение метастабильных структурных аномалий воды под действием электро-статического поля высокой напряженности у электродов (до 8 млн. вольт на см.), что не влияет на рН и ОВП, но выраженно изменяет биофизические свойства растворенных молекул и условия активации веществ в ходе ферментных реакций, изменяет реакции на границе пленок и клеточных мембран и пр.
В настоящее время имеется единственный патент на использование ЭХАС в клинической практике (Подколзин и соавт., 1993) для лечения гнойных ран, ожогов, воспалительных процессов слизистых оболочек и ряда иных патологических процессов кожи и слизистых.
Отмечены быстрая санация ран, раннее развитие грануляционной ткани, ускорение в 1,4 - 2 раза сроков заживления, улучшение приживления пересаживаемой кожи при ожогах, улучшение результатов лечения стоматологических и ЛОР-заболеваний и другие несомненные показатели благоприятного эффекта ЭХАС.
Отдельные клинические наблюдения, проведенные в других клинических группах, подтвердили эти наблюдения.
Так, показаны выраженные антибактериальные свойства анолита, превосходящие эффект многих современных антибиотиков, эффективность подавления роста таких устойчивых микроорганизмов, как синегнойная палочка, грибки кандида, другая устойчивая к антибиотикам микрофлора.
Католит способствовал ускорению регенерации, заживления и восстановления функций, снимал тканевой ацидоз и нормализовал многие параметры организма, нарушаемые в ходе патологического процесса.
Выраженные иммуностимулирующие свойства католита были показаны в прямых экспериментах с иммунизацией мышей эритроцитами барана. Количество антитело-образующих клеток (АТОК) в тесте Ерне на 4-й день после иммунизации животных повышалось в 4-6 раз. Введение католита позволяло восстановить иммунитет у мышей при моделировании иммунодефицита на примере фенилгидразиновой анемии, а также в случае экспериментального токсического гепатита, вызванного введением четырех-хлористого углерода, что позволяет использовать ЭХАС в случае вторичных иммунодефицитов.
Был исследован механизм иммуностимуляции в тестах кооперации Т-В лимфоцитов в системе Миллера и показано, что католит интенсифицирует процессы кооперации Т-В типа и одновременно угнетает функциональную активность антигенспецифических Т-супрессоров как на стадии формирования, так и отменяет их функциональный эффект для уже имеющихся клеток.
Важнейшим эффектом ЭХАС является способность метода очищать воду, используемую для самых различных целей.
Сравнение различных методов очистки воды показало высокие преимущества данного метода в сравнении со всеми известными способами очистки: микрофильтрации, ультрафильтрации, обратного осмоса, дистилляции, сорбции, ионного обмена, серебрения, иодирования, озонирования, хлорирования, ультрафиолетового облучения, кипячения, электродиализа, коагуляции, электролиза без диафрагмы и др.
Так, при 4-бальной системе оценки анолит обеспечивает 4-бальное очищение практически по всем параметрам: бактериального и вирусного загрязненяи, содержания микробных токсинов, фенолов, хлорорганических соединений, других органических загрязнителей, ионов тяжелых металлов и обеспечивается удовлетворительное удаление избытка минеральных солей.
Испытания показали, что анолит можно хранить и транспортировать в открытой и закрытой посуде без потери качества продукта в течение недель и месяцев; католит в нормальных условиях сохраняет свои качества в течение 1-2-х часов; в доверху плотно закупоренных бутылях - в течение 1-3-х дней.
Обращает на себя внимание, что ЭХАС - экологически чистые и совершенно не загрязняющие внешнюю среду жидкости - через несколько часов в окружающей среде они распадаются естественным образом, превращаясь в обычную воду, к тому же санируя среду обитания.
Установки типа "ИЗУМРУД" и "КРИСТАЛЛ" разрешены в настоящее время к производству для очистки питьевой воды, основные узлы запатентованы в СНГ и за рубежом и не имеют аналогов в мире.
Уже в настоящее время ЭХАС могут быть широко применены в медицине. Разрешено использование ЭХАС как экологически чистых моющих и дезинфицирующих растворов исходя из их уникальных бактериостатических свойств.
Выпускаются промышленно аппараты для очистки воды, в том числе для получения питьевой воды.
Имеется патентованный метод для наружного применения ЭХАС в медицине.
Наиболее перспективным представляется применение ЭХАС в медицине по следующим направлениям:
- использование АНОЛИТА как бактериостатического средства при инфицированных ранах, нагноительных внутренних процессах, воспалительных заболеваниях инфекционной природы самого различного происхождения и локализации (раны, ожоги, ЛОР- и стоматологические заболевания, кожные болезни, нефрология и пр.),
- использование КАТОЛИТА как средства, коррегирующего тканевой ацидоз, стимулирующего процессы регенерации тканей (раны, ожоги в фазе заживления, внутренние болезни),
- использование ЭХАС для коррекции острых (реанимационные состояния, острая фаза различных заболеваний) и хронических (болезни обмена, кожные болезни, болезни почек и печени и пр.) нарушения ОВП,
- использование ЭХАС как радиопротекторов при поражении ионизирующим облучением, лечебном воздействии ИО и цитостатиков,
- использование слабо минерализованных ЭХАС (водопроводная вода) получаемых на установках типа "Изумруд" или "Кристалл" как средства широкого общекоррегирующего и адаптогенного действия в ходе наружного (ванны, души) и внутреннего (для питья) применения,
Особый интерес представляет использование ЭХАС в геронтологической практике в связи с множественным благоприятным действием этих соединений на саму основу метаболических изменений при естественном и патологическом старении. Исходя из теории свободно-радикальных механизмов повреждения, представлений об энергетической недостаточности организма в пожилом возрасте и тканевом ацидозе и других, центральных для процесса старения механизмов, можно рекомендовать самое широкое применение ЭХАС для влияния на ключевые факторы старения и на вторичные его проявления, равно как и для повышения общего энергостатуса организма.
Использование ЭХАС для коррекции ОВП организма позволяет постоянно поддерживать энергостатус его на высоком уровне и противостоять самым различным внешним повреждающим воздействиям.
ВЛИЯНИЕ ЭХАС НА АНТИОКСИДАНТНЫЕ ФЕРМЕНТЫ
Мы исследовали влияние ЭХАС на активность всех трех основных ферментов антиоксидантной защиты организма по выше описанным методам: активность каталазы определяли в реакции перекиси с молибдатом аммония, активность пероксидазы – по реакции с индигокармином, активность СОД – по реакции ингибирования восстановления НСТ супероксидными ионами, генерируемыми в реакции НАД*Н с ФМС. В качестве стандартов мы использовали отмытые и лизированные эритроциты кролика, хранящиеся в замороженном состоянии до момента исследования.
ЭХАС получали пропуска раствор хлористого натрия (1 г на 1 Л) через установку "Изумруд", имеющую 2 реакционных блока и обеспечивающую в наших условиях получение ЭХАС с окислительно-восстановительным потенциалом от +200 мВ до – 70 мВ. Определение и постоянный контроль за ОВП в течение опыта проводился на рН-метре с платиновыми электродами в стандартном режиме определения ОВП проб.
ВЛИЯНИЕ ЭХАС НА АКТИВНОСТЬ КАТАЛАЗЫ ЭРИТРОЦИТОВ КРОЛИКА
Мы использовали 0,1 мл лизированных 1:1000 эритроцитов кролика, обеспечивающих снижение поглощения света при 410 нм (эквивалентно снижению содержания субстрата, доступного для молибдата аммония) на 50%.
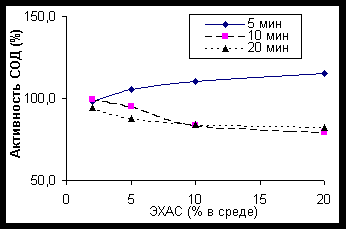
Рисунок 1. Влияние ЭХАС на каталазу эритроцитов кролика (время- и дозо-зависимые эффекты) Преинкубация пробы эритроцитов с различными концентрациями ЭХАС в течение 5, 10 и 20 минут показала (рисунок 1), что во всех случаях наблюдается двуфазный эффект ЭХАС: быстрое активирование фермента и последующее возвращение с прежнему уровню активности или слабое ингибирование.
В зависимости от ОВП ЭХАС эффект также достигал максимума, а затем снижался в зависимости от концентрации ЭХАС в среде (рисунок 2), 10 минут преинкубации с ЭХАС). Анализ данных позволяет в целом заключить, что с повышением ОВП и концентрации ЭХАС в среде снижается и необходимое для активации фермента время инкубации.
Рисунок 2. Влияние ЭХАС на каталазу эритроцитов кролика (Влияние ОВП и концентрации ЭХАС в среде инкубации) ВЛИЯНИЕ ЭХАС НА АКТИВНОСТЬ ПЕРОКСИДАЗЫ ЭРИТРОЦИТОВ КРОЛИКА
Изучение влияния ЭХАС на активность пероксидазы эритроцитов кролика показало, в целом, те же закономерности, что и влияние ЭХАС на активность каталазы. Для исследования влияния ЭХАС мы определили оптимальное время реакции, составившее в наших условиях 90-120 минут, достаточное для регистрации эффектов ЭХАС.
Титры перекиси и индигокармина также были оттитрованы в основной реакции, так как концентрации реактивов значимо влияли на точность определения реакции, регистрируемой по времени достижения стандартной окраски (в нашем случае – до полного обесцвечивания раствора, когда розовая окраска еще не начала развиваться), это обеспечило точность реакции в пределах 5-7 сек.
Чем больше концентрация ЭХАС использовалась для преинкубации с эритроцитами, тем меньшее время были необходимо для достижения максимума активации фермента (рисунок 3).
Рисунок 3. Влияние ЭХАС на активность пероксидазы эритроцитов кролика (время- и дозо- зависимость). Величина ОВП ЭХАС = - 50 мВ В отличие от каталазы, ясно наблюдались отсроченные влияния второй фазы – подавление активности пероксидазы при высоких концентрациях ЭХАС в среде и длительном времени влияния ЭХАС на лизированные эритроциты. Сходные данные отмечены и при исследовании эффекта ЭХАС с различным ОВП (рисунок 4).
Рисунок 4. Влияние ЭХАС на активность пероксидазы эритроцитов кролика (влияние ОВП) ВЛИЯНИЕ ЭХАС НА АКТИВНОСТЬ СУПЕРОКСИДДИСМУТАЗЫ
Изучение влияния ЭХАС на активность рекомбинантной СОД человека показало принципиально те же влияния, что и на активность каталазы и пероксидазы: в малых концентрациях, при небольших временах инкубации и при небольших значениях ОВП имеет место стимулирование фермента, тогда как повышение концентрации ЭХАС в среде, увеличение отрицательных значений ОВП и продление времени преинкубации ЭХАС с ферментом ведет к подавлению активности СОД (рисунки 5-6).
Аналогичное действие ЭХАС оказывали и на СОД эритроцитов кролика: преинкубация ЭХАС в течение 5 минут с лизатом эритроцитов стимулировала эффект СОД в широком диапазоне доз (но повышение дозы снижало эффект!), преикубация в течение 10 минут имела бифазный вид влияния, а в течение 20 минут – только ингибировала активность СОД.
Причина действия ЭХАС заключается, видимо, в том, что ЭХАС сами были способны генерировать продукцию супероксидных радикалов, оказывая, таким образом, умеренное тренирующее действие на активность ферментов антиокосидантной защиты организма. Наличие отрицательного ОВП для ЭХАС указывает на избыток электронов, возникающий при электрохимической активации, что, в принципе, как раз и является условием генерации в биологических системах ионов супероксида.
Рисунок 5. Эффект ЭХАС на активность рекомбинантной СОД (дозо-временная - зависимость)
Рисунок 6. Эффект ЭХАС на активность рекомбинантной СОД (ОВП - зависимость) Кроме того, большие отрицательные значения ОВП (-170 мВ в нашем случае) были способны инактивировать (видимо, окисляя) ингредиенты основной реакции: уже 5 минут инкубации с таким ЭХАС вызывало 22-25% снижение реакции ФМС+НАД*Н и НТС.
Таким образом, во всех случаях влияния ЭХАС на ферменты антиоксидантной защиты организма отмечены ясные дозо-зависимые влияния, зависимые от величины ОВП, концентрации ЭХАС в среде и времени преинкубации с эритроцитами. В целом, по любому параметру влияния ЭХАС достигали некоторого максимума и затем снижались, переходя в подавление активности фермента.
Влияние ЭХАС, видимо, связано с генерацией ЭХАС свободных радикалов, что в умеренных количествах способствует активации ферментов антиоксидантной защиты, то есть, ЭХАС оказывают тренирующее действие на активность ферментов антиоксидантной защиты. В больших количествах) дозах и длительном времени инкубации) ЭХАС подавляет активность этих ферментов, но в обычных условиях в организме практически не возможно достичь ингибирующих концентраций ЭХАС.
Таким образом, в зависимости от величины ОВП дозы и длительности действия, ЭХАС способны стимулировать антиоксидантные ферменты, генерировать продукцию свободных радикалов и инактивировать химические, и видимо биологические, молекулы (последний эффект лежит, очевидно, в основе известного антисептического действия ЭХАС при местном применении в значительных дозах, но фактические не достижим при применении вовнутрь).
Применение ЭХАС в клинической практике вполне оправдано для влияния на активность ферментов антиоксидантной системы организма и может объяснять многие влияния, отмеченные клиницистами при изучении влияния ЭХАС на различные физиологические и патологические процессы в организме как в клинической практике, так и в эксперименте на животных.
Так как антиоксидантные ферменты могут выражено влиять на явления старения клеток и тканей, а использование установок "Изумруд" одобрено для бытовой очистки воды, применение ЭХАС в широкой практике для практически здоровых лиц может быть эффективно тренирующим влиянием на ферменты антиоксидантной защиты организма, что дополняет экологическое влияние очистки воды и известные эффект активации типа "талой" воды – "гомогенизацию" воды (разрушение вторичной структуры воды) в ходе прохождения воды в таких установках через поля высокой электростатической активности.
ВЛИЯНИЕ ЭХАС НА АКТИВНОСТЬ ФЕРМЕНТОВ АНТИКОСИДАЗНОЙ ЗАЩИТЫ ОРГАНИЗМА in vivo
Для изучения влияния ЭХАС на активность ферментов антиоксидзной защиты в целостном организме исследовали влияние введения ЭХАС кроликам in vivo. Кролики получали с питьем в течение 2-х суток по 300 мл ЭХАС с начальной активностью ОВП= -70 мВ (содержание NaCl = 200 мг на 1 л). Затем исследовали уровень активности трех основных ферментов (пероксидазы, каталазы и СОД) в крови, а также реакцию этих ферментов на 10-минутную инкубацию со стандартным раствором ЭХАС (10% ЭХАС с 1 г на 1 л NaCl и с ОВП=-65 мВ) in vitro. Всего в контроле было 4 кролика и в опыте 10 кроликов.
ПЕРОКСИДАЗА.
Активность пероксидазы в контрольной группе составила в среднем 69,8 сек со стандартным отклонением 4,65 сек (разброс данных составил от 65 сек до 76 сек). В опытной группе время реакции снижалось, что указывает на активацию пероксидазы, до 52,9 сек (стандартное отклонение 17,99 сек), при более значительном разбросе активности фермента: от 30 сек до 75 сек. Видимо, такой разброс указывает на индивидуальные различия в активации фермента ЭХАС.
При воздействии ЭХАС на лизат эритроцитов оказалось, что в среднем имеет место тенденция к активации фермента в обеих группах, однако, большие разбросы данных делают заключение по средним значениям не достоверным (активация на 5,9 и 6,2 сек для контрольной и опытной групп соответственно при стандартном отклонении 11,5 и 8,9 сек).
Рассмотрение влияния ЭХАС по каждому из животных отдельно также не позволяет выявить ясных отличий – как при высоких, так и при низких значениях активности фермента влияние ЭХАС in vitro проявлялось как в активировании, так и в подавлении реакции.
КАТАЛАЗА
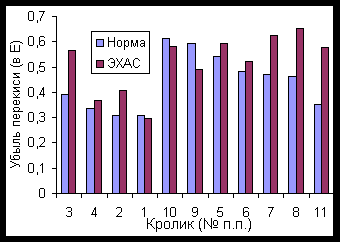
Активность каталазы в контрольной группе составила в среднем 0,335 условных единиц (выражено в единицах убыли экстинкции, что пропорцоионально количеству ферментированного субстрата – перекиси водорода), со стандартным отклонением 0,041 у.е. (разброс данных составил от 0,305 до 0,393 у.е.).
В опытной группе потребление субстрата за время реакции возрастало (наблюдалось активирование каталазы) до 0,503 у.е. (стандартное отклонение 0,089 у.е.), при более значительном разбросе активности фермента: от 0,353 до 0,613 у.е. Видимо, такой разброс указывает на индивидуальные различия в активации фермента под действием ЭХАС.
При воздействии ЭХАС на лизат эритроцитов оказалось, что в среднем имеет место тенденция к активации фермента в обеих группах (на 21,1 и 19,1 %% для контрольной и опытной групп соответственно), однако, большие разбросы данных делают заключение по средним значениям не достоверным (стандартные отклонения составили соответственно по группам 21,6 и 28,1 %%).
Рассмотрение влияния ЭХАС по каждому из животных отдельно показывает, что поение животных ЭХАС вызывает у всех (кроме кролика № 11) активацию фермента.
Сохраняется при средних и небольших значениях активации также стимулирующее влияние ЭХАС на лизат эритроцитов in vitro, которое хорошо видно для котнтрольных кроликов.
В то же время, сверхактивность (кролики 10 и 9) фермента in vivo сопровождается реверсией действия ЭХАС на лизат: в этом случае активность каталазы снижается (Рисунок 7).
СУПЕРОКСИДДИСМУТАЗА
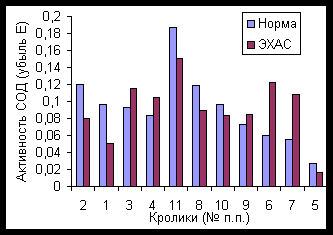
Активность СОД в контрольной группе составила в среднем 0,098 условных единиц (выражено в единицах убыли экстинкции, что пропорционально количеству НСТ, защищенного от перексида, генерированного в реакции НАД*Н с ФМС), со стандартным отклонением 0,016 у.е. (разброс данных составил от 0,265 до 0,302 у.е.).
Рисунок 7. Влияние ЭХАС на активацию каталазы кролика при введении in vivo
Активность каталазы выражена в убыли экстинкции (Е), что соответствует ферментированному субстрату. Повышению Е соответствует повышение активности фермента
В опытной группе имело место некоторое подавление активности СОД - до 0,088 у.е. (стандартное отклонение 0,053 у.е.), при более значительном разбросе активности фермента: от 0,198 до 0,358 у.е. Видимо, такой разброс указывает на индивидуальные различия в активации фермента под действием ЭХАС.
При воздействии ЭХАС на лизат эритроцитов оказалось, что в среднем имеет место тенденция к снижению активности СОД для контрольной группы (7,6%) при значительном стандартном отклонении (38,2 %), делающем заключение по средним не корректным, тогда как для опытной группы имело место активирование СОД (17,2%), но также при значительном стандартном отклонении (58,8%)
Рассмотрение влияния ЭХАС по каждому из животных отдельно показывает, что поение животных ЭХАС вызывает как повышение, так и снижение активности СОД (Рисунок 8), причем как для контроля, так и для опыта повышенные значения активности СОД сопровождаются реверсией реакции на ЭХАС в тесте in vitro, тогда как сниженные значения СОД – активацией на воздействие ЭХАС на выделенные эритроциты кролика. Исключение составил кролик № 5 с очень низкими значениями СОД, который реагировал на ЭХАС дальнейшим снижением активности СОД.
Рисунок 8. Влияние ЭХАС на активацию СОД кролика при введении in vivo
Активность СОД выражена в убыли экстинкции (Е), в присутствии СОД, что соответствует количеству защищенного от супероксида субстрата (НСТ). Повышению Е соответствует повышение активности фермента.
Таким образом, в целом, влияние ЭХАС на активность ферментов антиоксидантной защиты у животных in vivo имеет, в принципе, те же характерные черты, что и влияние в тестах in vitro, однако, примешивается влияние индивидуальной реактивности к ЭХАС, что, скорее всего, отражается на временнном типе реакции на введение ЭХАС – одни животные могут в тестированные сроки иметь фазу активации, тогда как другие – фазу компенсаторного снижения активности фермента после введения ЭХАС. На это указывают значительные индивидуальные разбросы активности фермента в опытной группе при достаточно однородных значениях в контроле и различия в чувствительности к ЭХАС в тесте in vit ro для животных с высокой и низкой активностью фермента.
Влияния ЭХАС на каталазу были более длительными, чем на ферменты защиты от пероксидных радикалов, что соответствует, в принципе, стратегии действия этих ферментов: СОД и пероксидаза инактивируют реакционные агенты с очень короткими сроками жизни, тогда как каталаза – со значительно более длительными сроками.
Особое значение имеет безусловное выраженное индивидуальное различие во влиянии ЭХАС на изучаемые ферменты.
В целом, можно описать действие ЭХАС на активность ферментов антиоксидазной защиты организма, как время- и дозо- зависимое быстрое активирование, сменяющееся фазой понижения активности, при выраженном индивиудальном различии в реактивности. Эффект ЭХАС скорее всего связан с прямым генерированием АФК (видимо, в результате избытка электронов при отрицательном ОВП), что оказывает тренирующее действие на систему ферментов и может быть зарегистировано прямо в эксперименте; кроме того, при сверхнизких (ниже - 200 мв) отрицательных значениях ОВП имеет место прямое химическое (окислительное) инактивирование химических и, видимо, биологических макромолекул.
Исходя из сказанного, можно считать показаниями к действию ЭХАС:
1. Азизова О.А.,Осипов А.Н., Савов В.М. и др. // Биофизика. 1985. Т. 30. С.36-39.
СИСТЕМА АНТИОКСИДАНТНОЙ ЗАЩИТЫ ОРГАНИЗМА И СТАРЕНИЕ
А.А. Подколзин, А.Г. Мегреладзе, В.И. Донцов, С.Д. Арутюнов, О.М. Мрикаева , Е.А. Жукова
Национальный Геронтологический Центр
Научно-исследовательский центр Московского государственного медико-стоматологического университета МЗ РФ
Основные механизмы появления АФК в организме связаны обычно с нарушениями функционирования электроннотранспортных цепей митохондрий или микросом, а также при изменении свойств дегидрогеназ. Особняком стоит нормальный процесс формирования АФК фагоцитами в ходе стимуляции неспецифической защиты организма.
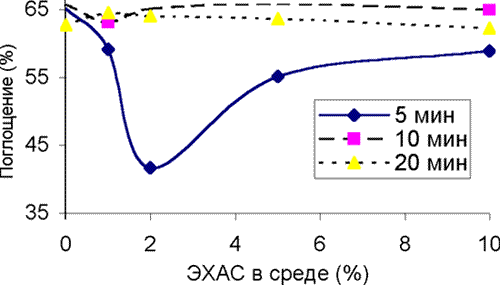
По оси ординат – поглощение субстрата (перекиси водорода + молибдат аммония)
Снижение показателей равноценной активированию каталазы
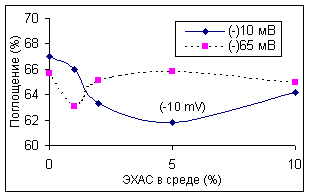
Обозначения как на рисунке 1
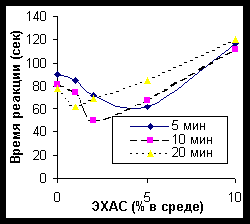
По оси ординат – время реакции. Снижение времени реакции указывает на активирование пероксидазы.
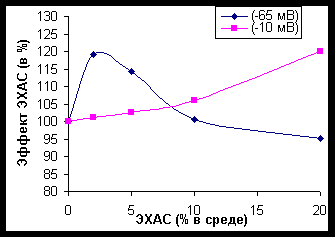
Время преинкубации – 10 минут
По оси ординат – эффект ЭХАС по отношению к контролю, принятому за 100%
По оси ординат – эффект ЭХАС в %
(контрольная активность СОД принята за 100%) ОВП = - 65 мВ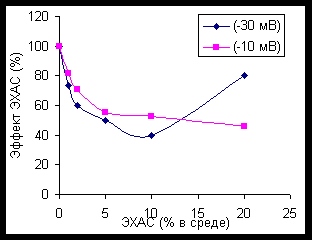
По оси ординат – эффект ЭХАС в %
(контрольная активность СОД принята за 100%)
ЛИТЕРАТУРА
2.Асатиани В.С. Ферментные методы анализа. М.:Наука. 1969. с.612
3. Аль-Хадиди М. Влияние ГБО, антиоксидантной терапии и их комбинации на свободнорадикальные процессы и клиническое течение стенокардии напряжения. Автореф. дис. канд. мед. наук. М.1987.
4. Анисимов В.Н.,Арутюнян А.В.,Опарина Т.И. и др. // Росс. физиол. журн. 1999. Вып.85. №4. С.502-507.
5. Бабенко А.Г., Гойницкий М.Н. // Лаб. дело. 1976. №3. С.157-158.
6. Бах А.Н., Зубкова С.А. // В: Сборник избранных трудов. П/ред. Бах А.Н. Л., 1997. С.412-415.
7. Бенина Н.Ф., Чегунова М.И. //Клин. Мед. 1974. №3. С.26-28.
8. Бобоходжаев М.Х. //Здравоох. Таджикистана. 1973. №6. С.11-15.
9. Ванин А.Ф. //Биохимия.1967.Т.32.С.277-282.
10. Владимиров Ю.А., Арчаков А.И. Перекисное окисление липидов в биомембранах. М.: Наука. 1972.
11. Второе республиканское совещание по электроактивированным средам. Казань. 1987
12. Гончарик И.И. //Актуальные проблемы теорет. и клинич. медицины. Минск. 1973. С.227-228.
13. Голубев А.Г.//Биохимия. 1996.Вып.61. С.2018-2039.
14. Громашевская Л.Л., Стремицкий Г.Ф., Гебеш В.В., Макаровская А.И. // Актуальные проблемы теорет. и клинич. медицины. Минск.1973. С.38-39.
15. Гродзинский Д.М., Войтенко В.П.,Кут- лахмедов Ю.А.,Кольтовер В.К. // Надежность и старение биологических систем. Киев: Наукова Думка, 1987.
16. Гуськова Р.А., Виленчик М.М., Кольтовер В.К. //Биофизика. 1998. Т.25. С.102-105.
17. Деев А.И., Осис Ю.Г., Формазюк В.Е. и др. //Биофизика. 1987. Т. 32. С. 629-636.
18. Донцов В.И., Крутько В.Н., Подколзин А.А. Старение: механизмы и пути преодоления. М.: Биоинформсервис. 1997.
19. ДубинаЕ.Е. и др.//Лаб.дело. 1988. N8. С.16-19.
20. Дупленко Ю.К. Старение: очерки развития проблемы. Л.: Наука. 1985.
21. Зенков Н.К., Меньшикова Е.Б.//Успехи современной биологии. 1993.Т.113.Вып.3. С.286-296.
22. Каган В.Е., Азизова О.А., Архипенко Ю.В. и др. //Биофизика. 1977. Т. 22. С. 625-630.
23. Каган В.Е., Сербинова А.Е.,Минин А.А. и др. //Биохимия. 1985. Т. 50. С. 986-991.
24. Клебанов Г.И., Туркменова Э.М., Крейнина М.В. и др.//Биол.мембраны.1987.Т.4. С. 1084-1092.
25. Красновский А.А. Синглетный кислород в фотобиотических процессах.:Дис.д-ра биол.наук.. М., 1983. 356 с.
26. Кольтовер В.К.// Успехи геронтологии. 1998. Вып.2. С.37-42.
27. Коган А.Х.//Вестник РАМН. 1999.№ 2.С.7-10.
28. Кольтовер В.К.//Успехи геронтологии. 1998. Вып.2. С.37-42.
29. Кольтовер В.К. // Хим.физика. 1996. Т.15. С.101-106.
30. Королюк М.А., Иванова Л.И.,Майорова И.Г., Токорева В.Е.//Лаб.дело. 1988. N1.С. 16-19.
31. Коган А.Х.//Вестник РАМН.1999.№ 2. С.7-10.
32. Коган А.Х., Кудрин А.Н., Лосев Н.И. Антиоксидантная защита сердца при экспериментальном инфаркте миокарда. М.:1987.
33. Коган А.Х., Кудрин А.Н.,Кактурский Л.В. и др.//Пат. физиол. 1992. №.2. С. 5-15.
34. Коновалова Г.Г., Лапкин В.З., Бескровнова Н.Н. //Арх.патол 1989. Т.51. №6. С.19-24.
35. Кульберг А.Я.,Елистратова И.А.,Тарханова И.А. и др. //Иммунология. 1986, №2. С.14-18.
36. Кульберг А.Я.,Петяев И.М.,Замотаева Н.Г.//Иммунология. 1988. №3. С.37-40.
37. Кульберг А.Я. Экологический кризис: стратегия выживания. М.:1994.
38. Лапкин В.З.,Коган А.Х.,Ковалевская А.Л. и др.// Бюл. эксперим. биол. мед. 1982. Т.93. №5. С.58-60.
39. Маянский А.Н., Маянский Д.Н. Очерки о нейтрофиле и макрофаге. Новосибирск. Наука. 1983. 264 с.
40. Меерсон Ф.З., Адыкалиев Н.А., Голубева Л.И. //Бюл. эксперим. биол. мед. 1981. Т. 91. С. 1167-1169.
41. Меньшикова Е.Б., Зенков Н.К.// Тер.архив. 1991. №11. С.85.
42. Осипов А.Н., Азизова О.А.,Владимиров Ю.В. Активные формы кислорода и их роль в организме.//Успехи.биол.химии. 1990. Т. 31. С. 180-208
43. Обухова Л.К., Эммануэль Н.М. // Успехи химии. 1983. Т.52. С.353-372.
44.Петяев И.М.,Кульберг А.Я.// Иммунология. 1988. №5. С.12-14.
45. Подколзин А.А., Донцов В.И. Факторы малой интенсивности в биоактивации и иммунокоррекции. М.:1995.
46. Подколзин А.А., Хасанов Р.Ш. Заявка на изобретение N 365284.13 (121800) от 9.08.1983.
47. Раппопорт Г.Л., Кекало Л.А., Бабенко Н.А. и др. //Пульмонология. Киев. 1977. Вып. 3. С.79-82.
48. Рябинский В.С. //Лаб.дело. 1970. № 3. С. 177-179.
49. Скулачев В.П//Биохимия. 1998. Т.63. Вып.11. С.1570-1579.
50. Тез. Всерос. конф."Методы и средства стерилизации и дезинфекции в медицине".М., 1992.
51. Тез. Второй Всерос. конф. Применение электроактивированных методов в медицине". Конаково. 1989.
52. Фролькис В.В., Мурадян Х.К. Экспери-ментальные пути продления жизни. //Л.:Наука. 1988.
53. Шукурова Р.А. //Здравоохр. Таджикистана. 1974. № 6. С. 31-33.
54. Чевари С., Чаба И., Секей Й. //Лаб.дело. 1981. N 11 С. 678-680.
55. Ченеев С.Б. //Вестник РАМН.1999.№2. С.10-15.
56. Ченеев С.Б., Кульберг А.Я.// Иммунология. 1995. №2. С.9-12.
57. Albich J.M., McCarthy C.A.,Hurst J.K. //Proc.Nat.Acad.Sci. USA 1981. Vol. 78. P. 210-216.
58. Allen R.C.//Chemical and biological generation of exited states.N.Y.:Acad.press. 1982. P.310—344.
59. Ambruso D.R., Johnston R.B.// J.Clin. Invest. 1981. Vol.67.P.350-360.
60. Anbar M.,Neta P.// Intern.J. Appl. Rediat. Isot. 1967. Vol.18,P.495-523.
61. Anisimov V.N.,Mylnikov S.V.,Khavinson V.Kh.//Mech.Ageing.Devel.1998.Vol.103.P.123-132.
62. Azbel M.Ya. // Proc.Natl.Acad.Sci.USA. 1994. Vol. 91. P.12453-12457.
63. Babior B.M. / /N.Eng.J.Med. 1978. Vol.298. P.659-668.
64. Babior B.M. // N.Eng.J.Med. 1978. Vol.298. P.721-725.
65. Bachur N.R., Gordon S.L., Ggee M.V.// Mol.Pharmacol. 1977. Vol.13, P.901 - 910.
66. Badway J.A.,Karnovsky M.L.//Annu.Rev. Biochem. 1980. Vol.49. P.695-716.
67. Bannister J.V.,Bannister W.H., Hill H.A., Thornalley P.J.//Biochem.et biophys. acta. 1982. Vol.715. P.116-120.
68. Banatti U., Morelli A.,Guida L.,de Flora A.//Biochem. and Biophys. Res. Commun. 1983. Vol. 111. P.980-987.
69. Bennett R.M., Eddie-Quartey A.C., Holt P.L.//Arthritis and Rheum. 1973. Vol.16. P.186-190.
70. Bennett R.M.,Skosey J.L.//Arthritis and Rheum.1977.Vol.20.P.84-90.
71. Bergmeyer H.U. / /Biochem.Ztschr. 1955. Vol.327. P.255-259.
72. Beyer W.F., Fridovich I.//Anal.Biochem. 1987. Vol.161. P.566-599.
73. Bhuyan K.C., Bhuyan D.K.//Biochim. et biophys.ata. 1977. vol.497. P.641-651.
74. Bielsky B.H., Arudi R.L.,Sutherland M.W.// J.Biol.Chem. 1983. Vol.258. P.4759-4761.
75. Bhunia A.K., Han H., Snowden A. etc.// J.Biol.Chem. 1998. Vol.272. P.15642-15649.
76. Bonnichsen R.K.//Acta chem.scand. 1948. Vol.2. P.781-787.
77. Bowern N., Ramshow I.A.,Clark I.A. etc.// J.Exp.Med. 1984. Vol.160.P.1532-1543.
78. Bolzan A.D., Brown O.A., Goya R.G., Bianchi M.S. //Exp.Gerontol.1995. Vol.30. P.169-175.
79. Bodnar A.G., Ouellette M., Frolkis M. etc.//Science. 1998. Vol. 279. P. 439-452.
80. Caruntu F.//Rom.Med.Rev. 1970.Vol.14.P.74-77.
81. Cameron D.J.//Jap.J.Med. 1986. Vol.56. N4. P.135-140.
82. Clark I., Hunt N.H.//Infect. and Immun. 1983. Vol.39. P.1-6.
83. Cohen G., DembiecD., Marcus J. // Anal. Biochem. 1970.Vol.34.P.30-38.
84. Cagnon M., Charbonneau R., Therrien J., Baril M.//Rev.canad.Biol.1976. Vol. 35. P.177-180.
85. Cutler R//Annu.N.Y.Acad.Sci. 1991. Vol.621. P.1-28.
86. Demopoulos H.B.,Flamm E.S.,Pietronigro D.D. etc.//Acta physiol. scand. 1982. Suppl.492. P.91-120.
87. Duran N. // Chemical and biological generation of exited states. N.Y.: Acad. press. 1982. P.345-359.
88. Dehaan J.B.,Cristiano F., Ianello R.C., Kola I. //Biochem.and Mol.Biol.Inter. 1995. Vol.35. P.1281-1297.
89. Dolle M.E.,Giese H.,Hopkins C.L. etc.// Nature Genetics. 1997.Vol.17.P.431-434.
90. Dogru-Abbasoglu S., Tanger-Toptani S., Ugernal B. etc. // Mech.Ageing.Devel. 1997. Vol.98. P.177-180.
91. Dypbukt J.M., Ankarkrona M., Burkitt M. etc.//J.Biol.Chem. 1994. Vol. 269. P. 30553-30560.
92. Edwards S.W., Hallett M.B., Lloyd D. etc.//FEBS Lett. 1983. Vol.161.P.60-64.
93. Flitter W.,Rawley D.A.,Haliwell B.//FEBS Lett. 1983. Vol.158. P.310-312.
94. Floyd R.A.//Arch. Biochem. and Biophys. 1983. Vol.225.P.263-270.
95. Frank B.//Angew.Chem.Intern. Ed. 1982. Vol.21. P.343-353.
95. Frew J.E., Jones P., Sholes G.// Anal. chim. acta. 1983. Vol.155. P.139-146.
97. Goth L.,Meszaros I., Nemeth H.//Clin. Chem. 1982. Vol. 28. P.1999-2000.
98. Goth L.//Hung.scient.instrum.1982.N 58.P.43-45.
99. Goth L., Nemeth H., Meszaros I.//Clin.Chem. 1983. Vol. 29. P. 741-743.
100. Gross J.,Hartwig A.,Golding A.//Z.med. Labortech. 1975. Bd. 16, S.336-339.
101. Gabing T.G.,Babior B.M.//Blood. 1979. Vol.53. N6. P.1133-1139.
102. Harman D. // Annu. N.Y. Acad.Sci. 1994. Vol.717. P.1-15.
103. Hejnecke J.W.//Free Rad.Biol.Med. 1987. Vol.3. P.65-76.
104. Hagen T.M., Yowe D.L., Bartholemew J.C. et al.//Proc.Natl.Acad.Sci.USA. 1997. Vol. 94. P.3064-3069.
105. Harman D. // Proc. Natl. Acad. Sci. USA. 1991. Vol. 88. P.5360-5363.
106. Ho Y.-S.,Magnenat J.-L.,Gargano M. etc.// Environ.Health Perspect.1998. Vol.106. P.1219-1228.
107. Ioshikawa Y., Yokoo T., Kitamura M. // Biochem. Biophys. Res. Commun. 1997. Vol. 240. P. 496-501.
108. Jacobs A. // Semin.Hematol. 1977. Vol. 14. P.89-113.
109. Keston A.S., Brandt R. // Anal. Biochem. 1965. Vol.11. P.1-9.
110. Khan A.U.//Science.1970. Vol.168. P.476-482.
111. Klebanoff S.J.,Clark R.A. The niutrophil: function and clinical disorders. Amsterdam: North-Holland. 1978. 313 p.
112. Kellog E.W., Fridowich I. // J.Gerontol. 1976. Vol.31. P.405-408.
113. Koltover V.K. //Age.1995.Vol.18. N.3. P.85-89.
114. Kroemer G. // Cell Death and Differentiation. 1997. Vol.4. P.443-456.
115. Kogan A.Kh.,Kudrin A.N.//J. Molec. Cell. Cardiol. 1980. Vol. 12. Suppl.1. P.75.
116. Kogan A.Kh., Grachyov S.,Sumarokov A.// Oxidativ Stress and Redox Regulation: Abstracts. Paris. 1996. P.300.
117. Lee C.M., Weindruch R., Aiken J.M. // Free Radical Biol. Med. 1997. Vol.22. P.1259-1269.
118. Luis D.R., Ortega M.G.,Lopez A.L., Gorge J.L.//Analyt.Biochem. 1977. Vol.80. P.409-415.
119. Lee S-L., Wang W.W., Fanburg B.L.//Free Rad.Biol.Med. 1998. Vol. 24. P. 855-858.
120. Marcuse R.,Johnston L.//J.Amer.Oil Chem. soc. 1973. Vol.50. P.387-391.
121. Massey V.//Biochim. et biophys.acta. 1959. Vol.34. P.255-262.
122. Matsuo M., Gomi F., Kuramoto K., Sagai M. //J.Gerontol. 1993. Vol.48. B.133-138.
123. McCord J.M.//Agents and Actions. 1980. Vol.10. P.522-527.
124. McCord J.M.,Fridovich I.//Agents and Actions. 1969. Vol.244. P.6049-6055.
125. Meerhof L.J., Roos D.//J.Reticuloendothel. Soc. 1980. Vol. 28. P.419-425.
126. Mello Filho A.C., Hoffman M.E., Meneghini R. //Biochem. J. 1984. Vol.218. P.273-275.
127. Mercel P.B.,Kearns D.R.//J/Amer.Chem. Soc. 1972. Vol.94. P.7244-7248.
128. Misra H.P., Frodowich I. //J.Biol.Chem. 1972. Vol.247. P.3170-3175.
129. Mustafa M.G. // Free Radicals Biol. and Med. 1990. Vol. 9. P.245.
130. Munakata T.,Semba U.,Shibuya Y.etc.// J.Immunol. 1985. Vol.134. N4. P.2449-2455.
131. Myers C.E.,McGuire W.P., Liss R.H. etc.// Science. 1977. Vol.197. P.165-167.
132. Nashio E., Watanabe Y.//Biochem. Biophys. Res. Commun. 1992. Vol. 232. P. 1-4
133. Nohl H.//Free radicals, aging and degenerative disease. N.Y.: Liss, 1986. P.77-97.
134. Nuttal S.L.,Martin U.,Hutchin T.etc.// Age&Ageing. 1998. Vol.27. P.34.
135. O`Connel M.J., Garner A.//Intern.J. Radiat. Biol. 1983. Vol.44. P.615-625.
136. Orr W.C., Sohal R.S. //Science. 1994. Vol.18. P. 127-137.
137. Ozawa T. //Physiol. Rev. 1997. Vol.77. N.2. P.425-464.
138. Papa S.,Skulachev V.P.//Molec.Cell. Biochem. 1997. Vol.174.P.305-319.
139. Perschke H., Broda E.//Nature. 1961. Vol.190. P.257-261.
140. Rao P.S.,Mueller H.S.//Advanc.Exp.Med. Biol. 1983. Vol.161. P.347-364.
141. Romero F.J.,Bosch-Morell F.,Romero M.J. etc.//Environ. Health Perspect. 1998. Vol.106. P.1229-1234.
142. Romson J., Hook B., Kunkel S. etc.// Circulation. 1983. Vol.67. P.1016-1023.
143. Rosen G.M., Finkelstein E., Rauckman E.J.//Arch.Biochem. and Biophys. 1982. Vol.215. P.367-379.
144. Roschupkin D.I., Pelenitsin A.B., Potapenko A. Ya. etc. // Photochem.and Photobiol. 1975. Vol.21. P.63-69.
145. Rowley D.A., Halliwell B. // FEBS Lett. 1982. Vol.142. P.39-41.
146 banyi C.M. //Free Radicals Biol. and Med. 1988. Vol. 4. P.107.
147. Saito H.,Hayakawa T.,Yui Y. etc.// Int.Arch.Allergy. 1987. Vol. 76. N1. P.92-96.
148. Segal A.W.,Jones O.T.G.//Nature.1978. Vol.276. P.515-518.
149. Sies H.//Amer. J. Med. 1991. Vol. 91. P. 31.
150. Sinha G.K., Mimnough E.G.// //Free Radicals Biol. and Med. 1990. Vol.8. P.567.
151. Shigenaga M.K., Hogen T.M., Ames B.N.// Proc..Nat.Acad.Sci.USA 1994. Vol.91. P.10771.
152. Suthanthiran M., Solomon S.D., Williams P.S. etc.//Nature. 1984.Vol.307. P.276-278.
153. Thomspson J.A., Hess M.L.//Progr. Cardiovasc. Dis. 1986. Vol.28. P.449-462.
154. Van Kessel K.P., Visser M.R., VanStrijp J.A. etc.//Immunology. 1987. Vol.62. N4. P.675.
155. Vladimirov Yu.A.//Free radicals, aging and degenerative diseases. N.Y.: liss, 1986. P.141-195.
156. Vladimirov Yu.A.//Sov.Med.Rev.B. Phy-sicochem Asp.Med. Harwood Acad.Publ. 1987. Vol.1. P.51-127.
157. Vladimirov Yu.A., Olenev V.I., Suslova N.B.,Cheremissina Z.P.//Adv.Lipid.Res. 1980. Vol.17. P.173-249.
158. Von Zglinicki T., Sarezki G., Doccke W. etc.//Exp.Cell.Res.1995.Vol. 220. P.186-193.
159. Ward P.A., Till G.O., Kunkel R. etc.// J.Clin.Invest. 1983. Vol.72. P.789-801.
160. Werns S.W., Shea M.J., Driscoll E.M. etc.// Circulat.Res. 1985. Vol.56. P.895-899.
161. White B.C., Aust S.D., Arfors K.E. atc.// Ann.Emerg.Med. 1984. Vol.13. pt 2. P.127-132.
162. Woodward B., Zabaria M. // J.Mol.and Cell.Cardiol. 1985. Vol.17. P.485-493.
163. Yan L.-J., Levine R.L., Sohal R.S.// Proc..Nat.Acad.Sci.USA. 1997. Vol.94. P.11168.
164. Yunis J.J.//Science. 1983. Vol.221. P.227.
Январь 2001 г.